牙周膜和牙齦干細胞實現肌腱組織再生的方法與流程
本發明涉及干細胞技術領域,尤其是涉及一種裝載tgf-β3、rgd修飾的并載入了牙周膜干細胞和牙齦間充質干細胞的海藻酸鹽微球共載體系的牙周膜和牙齦干細胞實現肌腱組織再生的方法。
背景技術:
肌腱損傷可引起患者長期疼痛、不適和功能障礙。其有限的自我修復能力使得用再生治療方法來功能性修復肌腱損傷顯得尤為必要。間充質干細胞的應用可能為肌腱再生和修復提供新的治療選擇。
因為取材方便,在牙源性間充質干細胞中的牙周膜干細胞和牙齦間充質干細胞引來廣泛關注,其可從口腔臨床中廢棄的生理組織中提取。該牙源性干細胞已經通過體內外的實驗證明了其多向分化的能力。另外,因為牙周韌帶含有肌腱樣的膠原束,可以承受口腔咬合力,在肌腱再生治療中,牙周膜干細胞表現出作為種子干細胞獨特的潛力。
研究表明tgf-β通路在肌腱再生中起著很重要的作用。tgf-β3由于它是一個與促進皮膚傷口和肌腱愈合無纖維疤痕形成有關的同種型。由tgf-β3引導的間充質干細胞分化,能促進肌腱組織形成。為了修飾微環境特性和引導細胞向特定表型分化,將海藻酸凝膠和rgd相結合。海藻酸鹽能提供一種利于mscs的空間分布的三維支架,從而能得到一種類似于體內微環境的結構組織。
最近,本研究團隊通過體內外實驗發現,結合rgd的海藻酸凝膠可用于復合pdlscs和gmscs用作軟骨再生,開發了一個能給牙源性mscs(pdlscs和gmscs)的復合提供三維架構的共載體系。假設裝有tgf-β3,復合pdlscs 和gmscs的rgd修飾的海藻酸水凝膠微球具有肌腱再生能力,對肌腱組織工程是一項有前景的應用。
技術實現要素:
本發明的目的在于提供一種牙周膜和牙齦干細胞實現肌腱組織再生的方法,為實現上述目的,本發明提供如下技術方案:
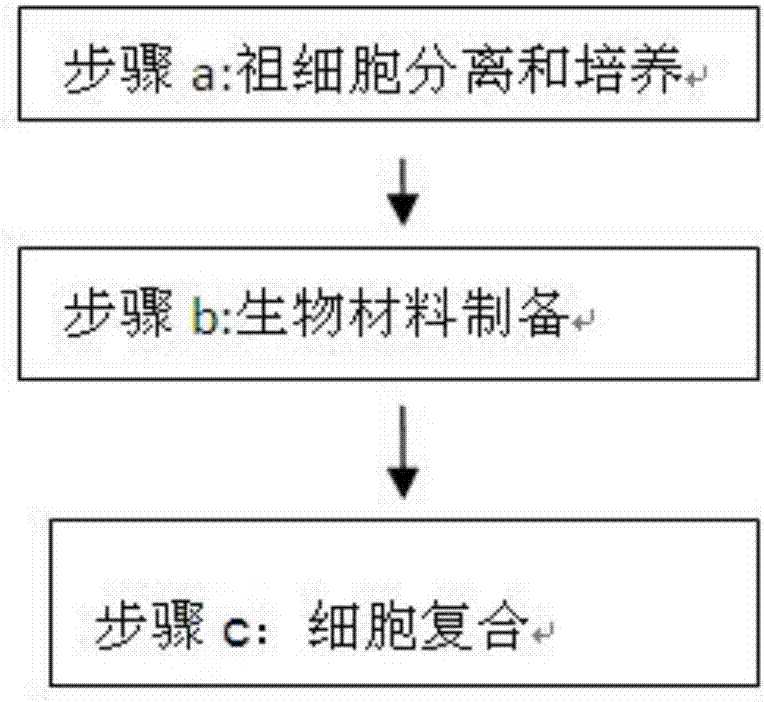
一種牙周膜和牙齦干細胞實現肌腱組織再生的方法,其特征在于:該方法包括以下步驟:
步驟a.祖細胞分離和培養
選擇并收集18-25歲因需拔牙的健康患者第三周磨牙的牙齦組織和牙齒,要求無任何牙周病史,送至實驗室;
步驟b.生物材料制備
選取rgd-coupled海藻酸鹽作為支架材料,對其進行處理以提高降解性,并與tgf-β3混合。
步驟c.細胞復合
將上述混合有tgf-β3配體的海藻酸鹽中復合牙源性干細胞,將其添加到cacl2溶液中以形成微球。
所述步驟b中還包括以下分步驟:
分步驟1,選取rgd-coupled的海藻酸鹽,對其進行純化和部分氧化(2%),以增加其降解性;
分步驟2,收集該海藻酸鹽、對其進行減壓、冷凍及干燥處理;
分步驟3,將上述經過處理后的海藻酸鹽與tfg-β3進行混合。
所述步驟c中還包括以下分步驟:
分步驟1,將步驟b中所得到的含有tgf-β3的rgd-coupled海藻酸鹽溶液分別與pdlscs、gmsc兩種細胞以2*106cells/ml的比例混合;
分步驟2,逐滴將上述混合物(msc-混合物)添加到100mmcacl2的溶液中行成微球;
分步驟3,將上述微球在37℃環境中孵育45分鐘以完成交聯;
分步驟4,用不完全的dmem清洗上述微球,清洗三遍。
本發明所提供的方法有益效果是:通過含有tgf-β3配體的rgd三肽耦合海藻酸鹽微球作為牙源性msc的復合載體,改善了牙源性細胞分化成肌腱樣組織的微環境,為肌腱組織工程提供了一個獨特的3d細胞交架。gmscs很容易從口腔中獲取,或者從廢棄組織中提取,這些msc源可以被認為是理想的干細胞庫來源,在基于msc的組織再生中很有潛力,該方法對于優化肌腱再生以及四肢骨骼修復等方面都有潛在應用前景。
附圖說明
圖1是本發明的步驟流程示意圖。
具體實施方式
下面將結合本發明實施例,對本發明實施例中的技術方案進行清楚、完整地描述,顯然,所描述的實施例僅僅是本發明一部分實施例,而不是全部的實施例。基于本發明中的實施例,本領域普通技術人員在沒有做出創造性勞動前提下所獲得的所有其他實施例,都屬于本發明保護的范圍。
實施例:參照圖所示,本發明所提供的一種牙周膜和牙齦干細胞實現肌腱組織再生的方法包括以下步驟:
步驟1.祖細胞分離和培養
牙齦組織和牙齒是從二十歲左右的來自南加州大學的健康男性患者的第三磨牙中收集的(18-25歲),無任何牙周病史的樣本才能被納入本研究。
步驟2.生物材料制備和細胞復合
使用rgd-coupled海藻酸鹽(novamatrixfmcbiopolymer,norway)作為支架材料。進行海藻酸的純化和部分氧化(2%)來增加其降解性。隨后,海藻酸鹽被收集起來,在減壓下冷凍干燥,和tgf-β3(invitrogen,carlsbad,ca)混合。
pdlscs、gmsc和hbmmscs(作為陽性對照)分別復合在含有tgf-β3配體的海藻酸鹽中。細胞被復合在海藻酸鹽的密度是2*106cells/ml。逐滴把msc-海藻酸鹽混合物添加到100mmcacl2溶液中形成微球。形成的微球在37℃孵育45分鐘來完成交聯,然后用不完全dmem洗三遍。沒有細胞(也不帶有tgf-β3)的rgd-coupled海藻酸鹽水凝膠作為陰性對照。
步驟3.活細胞/死細胞染色
培養14天后,復合的msc細胞的活性使用鈣黃綠素染色活細胞和溴化乙錠二聚體染色死細胞(invitrogen)。用nihimagej測量軟件(nih,bethesda,md)測量活細胞的百分比。通過沒有rgd的復合牙源性msc,hbmmscs的藻酸鹽微球培養14天后的活細胞/死細胞染色測定,來評估rgd三肽的存在對復合msc的活性的影響。
步驟4.掃描電鏡(sem)
通過掃描電子顯微鏡(sem)(jeol5300,peabody,ma)來觀察微球的形態和結構特征。在常規培養基培養14天后,msc藻酸鹽微球用2毫升pbs洗一遍,用1%戊二醛固定過夜。樣本使用酒精溶液分級脫水,濺射鍍黃金,并用掃描電鏡觀察。
步驟5.共聚焦激光掃描顯微鏡(clsm)
使用clsm(fluoviewfv10i,olympuscorp,tokyo,japan)進一步來觀察復合msc的藻酸鹽微球樣品形貌。在培養的第14天,復合的細胞用多聚甲醛(4%)固定30分鐘,用pbs(ph=7.4)清洗,然后用0.1%tritonx-100在室溫 下孵育5分鐘。抗cd146-fitc和cd105-pe抗體(abcam)作為msc的陽性標記物。此外,抗cd34-pe抗體(santacruzbiotechnology,inc.,santacruz,ca)作為陰性標記物。所有的標本用dapi進行細胞核染色復染。
步驟6.體外釋放曲線
為了描述tgf-β3、rgd-coupled海藻酸微球的釋放曲線,裝載三個不同濃度(10、50、100毫克/毫升)tgf-β3,在500ulhigh-glucosedmem與100u/ml青霉素和100ug/ml鏈霉素在48孔板中培養,37℃搖床一周。在每個選定的時間間隔(1、3、5、7、10、12、14天)收集其培養基,使用tgf-β3elisa試劑盒(r&dsystemsinc.,minneapolis,mn)來分析tgf-β3的釋放。在釋放曲線研究的最后,將余留的tgf-β3從微球中提取出來,通過50mm檸檬酸鈉脫水溶液(sigmaaldrich,stlouis,mo)在蒸餾水中溶解藻酸鹽支架,來測量總共釋放的tgf-β3的百分比。最后,釋放的tgf-β3的百分比通過兩周后的釋放量占最初添加量的百分比來計算。
步驟7.體外培養和免疫熒光染色
為了誘導腱向分化、復合的pdlscs和gmsc,hbmmscs(2*106cells/mltgf-β3裝載的海藻酸微球)在含有dmem15%的血清,2mml-glutamine,100nmdex,1003mascorbic酸2mmsodium丙酮酸(r&dsystemsinc.),100u/ml青霉素,100ug/ml鏈霉素的成腱培養基中培養。無細胞rgd-coupled沒有裝載tgf-β3海藻酸微球作為陰性對照。
誘導4周后,樣本用4%pfa固定,石蠟包埋切片。切片使用一抗tenomodulin(tnmd)eya1,eya2(fromsantacruzbiotechnology,inc.,dallas,tx),和scleraxis(abcam)標記4℃過夜,alexafluor染料標記的二抗(1:200稀釋;invitrogen)檢測,再用dapi復染。
步驟8.rna提取、逆轉錄和實時定量pcr
按照以往文獻發表的方法將細胞培養2周后,提取rna。10個藻酸鹽微球被收集起來,在一個包含50mm檸檬酸鈉脫水溶液和80mm氯化鈉(sigmaaldrich)的無菌解聚緩沖液溫和攪拌15-20分鐘使其溶解。溶散后,無莢膜的細胞在9400g離心機離心10分鐘,微球用pbs清洗,9400g再次離心3分鐘。總rna使用試劑盒試劑(invitrogen)提取。100ng總rna使用一個標注著iiicdna合成試劑盒(invitrogen)來合成單鏈的cdna。用2ddct方法進行數據分析,規范化看家基因gapdh(甘油醛3脫氫酶甘油醛)的ct。
步驟9.免疫印跡分析
培養四周后,干細胞的分化使用免疫印跡法進行了分析。干細胞藻酸鹽在檸檬酸緩沖液(6%w/v,ph值7.4)溶解。離心分離得到的細胞球,用pbs洗兩次,用蛋白質提取緩沖液(bio-rad,irvine,ca)溶解30s。離心分離棄上清,通過一個已知bsa標準系列濃度在一個數量級連續稀釋外推的bca試驗(pierce,rockford,il)測蛋白濃度。取相同質量的細胞裂解液,上樣,電泳,轉膜封閉。硝化纖維膜用針對孵化腱調蛋白(tnmd)(santacruzbiotechnology,inc.)的兔抗體孵育。免疫蛋白復合物用1:500(polyclonalgoatanti-rabbitigg/hrp,emdmillipore,billerica,ma)二抗檢測。剪膜,重新用抗b-actin的抗體檢測,以保證每一條帶添加的質量相同。化學熒光試劑(amershamlifescience,pittsburgh,pa)添加到膜上1分鐘,x光片自動洗片機來洗片,曝光不同時間來觀察圖像。
步驟10.肌腱組織再生動物模型
我們體內試驗中所有的動物均根據南加州大學對動物的使用和護理指導法規進行處理。pdlscs、gmsc和hbmmscs(大約4*106細胞)復合在裝滿tgf-β3的藻酸鹽微球上,10微球(500毫升)移植入5個月大的裸小鼠(harlan,livermore,ca;n1/44foreachgroup)背部皮下。8周后,小鼠被處死,微球和周圍組織利用組織學和免疫組織化學切片染色分析。無細胞的沒有tgf-β3的 rgd-coupled海藻酸微球作為陰性對照。
步驟11.組織學和免疫組織化學分析
組織學分析:標本用10%福爾馬林溶液固定,在一系列上升濃度的乙醇中脫水,石蠟包埋。用切片機切成6um切片,放在載玻片上。每個移植樣本隨機選擇4個橫切片,進行he和masson染色。另外,組織切片用偏振光顯微鏡觀察評價膠原合成。
免疫組化分析:石蠟切片浸泡在3%過氧化氫/甲醇15分鐘,抑制非特異性內源性過氧化物酶活性。切片用一抗(1:200-1:300稀釋)孵育1h。切片用anti-tenomodulin(santacruzbiotechnology,inc.,1:100dilution),anti-eya1,anti-eya2(santacruzbiotechnology,inc.,1:100dilution),anti-scleraxis(abcam,1:100dilution)進行免疫組織化學分析,并用蘇木精復染。
rgd三肽對復合msc肌腱再生能力的的影響與無rgd的藻酸鹽微球相比較。將微球用上述方法進行移植,并將切片進行he和masson染色。陽性染色由每個實驗組4個獨立樣本測定。從每個樣本隨機選擇五個區域,然后每一區域陽性染色區用nihimagej軟件測量,計算其占總區域的百分比。
步驟12.統計分析
用kruskal-wallisranksumtest來分析數據,a=0.05為顯著性檢驗水平。同時,如需要時,用two-tailedstudent’st-test來進行配對檢驗。定量數據表示為mean±sd。
結果分析:
1.生物材料制備和特性
用一個復合rgd海藻酸鹽體系來研究pdlscs和gmcss對肌腱組織分化和再生的能力。通過hbmmscs復合藻酸鹽微球作為陽性對照,而藻酸鹽微球作為陰性對照。此外還檢測了rgd-coupled海藻酸對msc的活性和分化作用。制造的藻酸鹽微球的平均直徑為1±0.1mm。pdlsc、gmsc和hbmmsc 組之間的活細胞比例沒有明顯的差異(p>0.05)。
經過兩周的培養,進一步用clsm檢測復合的牙源性干細胞的特性,顯示細胞表達干細胞特異性抗體cd146和cd105,但不表達cd34。這些結果證明了rgd耦合的海藻酸鹽作為pdlscs,gmsc以及hbmmscs復合的生物材料的適用性。
下一步,預備三種不同tgf-β3濃度(10、50和100毫克/毫升)的藻酸鹽微球,描繪兩周內tgf-β3從rgd耦合海藻酸支架的釋放曲線。總的釋放曲線證實tgf-β3釋放長達14天。通過評估最初tgf-β3占配體的比例,證實50毫克/毫升tgfb3呈現出最好的釋放曲線。因此接下來的實驗研究選擇了該濃度。
2.牙源性msc在體外的腱向分化
腱調蛋白(tnmd)是ii型跨膜糖蛋白,主要表達于肌腱和韌帶等組織。已經被證實早期表達scleraxis的祖細胞最終形成肌腱組織。eya1和eya2基因在肢體肌腱發育和編碼激活轉錄功能中被表達。這些轉錄因子參與包括肌腱和韌帶發育的許多細胞和發育過程。
對tgf-β3裝載的海藻酸鹽微球在肌腱再生中的分子機制進行研究。用幾個與肌腱分化相關的基因標記物來檢測相關基因的表達,包括轉錄因子scleraxis(scx),細胞外基質蛋白decorin(dcn)和表面標記tenomodulin(tnmd),以及二聚糖(bgy),這是一個一型的富含亮氨酸的蛋白多糖(slrp),參與調節膠原蛋白的原纖維形成。這些基因的表達水平是通過rt-pcr來證實和比較。然而對所有的檢測基因,pdlscs比gmsc或hbmmscs有顯著更高的表達水平(p<0.05)。在scx、dcntnmd或bgy表達水平上,hbmmscs和gmsc之間沒有的顯著差異(p>0.05)。然而,對于所有的測試基因,hbmmscs比gmsc有更高的表達。進一步的分析清晰地證實在細胞腱向分化中,tgf-β3比rgd扮演著更重要的角色。在tgf-β3配體存在下,對體外腱向分化兩周 的沒有支架復合的hbmmscspdlscs,gmsc的dcn和tnmd表達水平進行檢測。
3.體內異位肌腱再生
通過免疫缺陷的老鼠皮下異位移植實驗評估pdlscs和gmsc復合在rgd耦合的裝載tgf-β3的藻酸鹽微球中再生肌腱組織的能力。組織學切片he和masson染色顯示了波形排列的腱樣結構的纖維存在。實驗和陽性對照組三色染色進一步證實了移植8周后肌腱樣組織的存在,但在陰性對照組,無細胞移植組,沒有組織學證據顯示肌腱樣組織再生。pdlsc組比hbmmsc,gmsc組觀察到更多的黃色(網狀)染色的膠原。用免疫組化來檢測基因表達蛋白從而進一步描述牙源性msc介導的肌腱體內再生的特性。
- 還沒有人留言評論。精彩留言會獲得點贊!