用于治療慢性阻塞性肺病和哮喘的組合物和方法與流程
相關申請的交叉引用
本申請根據35u.s.c.119(e)要求于2009年4月22日提交的美國臨時專利申請第61/171,705號和于2009年12月2日提交的美國臨時專利申請第61/266,048號的權益,通過引用將兩篇專利申請全文并入本文。
關于序列表的聲明
以文本格式代替紙質版提供本申請相關的序列表,并且在此通過引用并入本說明書。包含序列表的文本文件命名為480266_404pc_sequence_listing.txt。該文本文件為67kb,創建于2010年4月22日,并且在提交本說明書同時,通過efs-web以電子方式提交了該文本文件。
發明背景
發明領域
本發明涉及用于治療慢性阻塞性肺病(copd)和哮喘的方法,具體而言,本發明涉及通過給予v型膠原(colv)或其耐受原性片段來治療copd和哮喘。
相關技術的描述
慢性阻塞性肺病(copd)是一類其中氣道變窄的肺部疾病。這導致進出肺部的氣流受限,引起氣短。與哮喘不同,氣流受限可逆性較差,并通常隨時間持續惡化。copd也被稱為慢性阻塞性肺部疾病(cold)、慢性阻塞性氣道疾病(coad)、慢性氣流通氣受限(cal)和慢性阻塞性呼吸道疾病。術語“copd”包括兩種主要的疾病狀態—肺氣腫和慢性阻塞性支氣管炎。
肺氣腫中,很多肺泡之間的壁被損傷,導致它們喪失其形狀并變得松軟。這種損傷還能破壞肺泡壁,導致更少和更大的肺泡替代很多微小的肺泡。
慢性阻塞性支氣管炎中,氣道內壁時常發炎和紅腫。這導致內壁增厚。在氣道中形成很多厚的黏液,使其呼吸艱難。
大多數患有copd的人既患肺氣腫又患慢性阻塞性支氣管炎。因此,通用術語“copd”是更精確的。
慢性支氣管炎和肺氣腫最常由吸煙引起;大約90%患有copd的患者目前或者過去是吸煙者。盡管大約50%的吸煙者發展為慢性支氣管炎,但是僅有15%的吸煙者發展為不可逆的氣道阻塞。某些其它哺乳動物,尤其是馬同樣會患copd。
與copd相關的氣道阻塞是進行性的,可以伴隨氣道高反應性,并且可以是部分可逆的。非特異的氣道高反應還可能在copd的發展中起作用,并且可以預示吸煙者肺部功能的衰退速率加速。
copd是死亡和傷殘的主要原因。當前其是美國和歐洲的第四大死亡原因。治療方針倡導早期檢測并執行戒煙行動以幫助降低由該疾病導致的發病率和死亡率。然而,由于諸多原因,早期檢測和診斷很難。
copd的發展需要花費數年,并且吸煙者通常否認由吸煙引起的任何有害效應,將呼吸困難增強的早期警告信號歸因于年齡信號。同樣,支氣管炎的急性發作通常不被全科醫生認為是copd的早期信號。很多患者表現出多于一種疾病(例如,慢性支氣管炎或哮喘性支氣管炎)的特征,使得精確診斷成為一種挑戰,尤其是在早期疾病時。同樣,很多患者并不尋求醫療幫助,直到他們遭受更嚴重的與肺功能減弱相關的癥狀時,例如呼吸困難、持續咳嗽和產生痰。因此,絕大多數患者沒有被診斷或治療直到他們處于更晚期的疾病階段時。
哮喘是折磨數百萬人群的氣道異質性障礙。氣道炎癥、高反應性和梗阻可以表征該疾病狀態。該疾病通常引起支氣管平滑肌系統的痙攣,并且影響上呼吸道和下呼吸道。有數種形式的哮喘,特征為嚴重程度不同。例如,輕度哮喘被界定為哮鳴的短暫發作,具有或不具有呼吸困難或咳嗽。中度哮喘被界定為哮鳴和呼吸困難,并且可以具有或不具有咳嗽和咳痰,但是通常會干擾日常活動和/或睡眠。重度哮喘的特征為由于呼吸困難導致能力喪失,并且受影響的患者通常無法正常進食或睡眠,極其焦慮并且經常會疲憊不堪。被稱為哮喘持續狀態的疾病狀態是最嚴重的哮喘形式,通常需要加強住院治療,并且甚至會致命。該疾病的發生可能是由于過敏性和非過敏性兩種機制。
盡管有數種可用來緩解與哮喘相關的癥狀和不適的治療,但是都沒有治愈。此外,目前的治療通常會引起副作用,使不適加重并發生其他致人虛弱的疾病狀態。輕度哮喘通常用β-腎上腺素藥以及抗組胺藥來治療,尤其對于兒童而言,從而預防或抑制間歇性發作。中度和重度哮喘通常用腎上腺素藥劑和支氣管擴張藥以及皮質類固醇來治療。由抗哮喘性藥劑導致的其它作用限制了它們的廣泛應用,所述其它影響包括頭痛、疲勞、口干、神經質以及某些情況下的成癮和物質濫用。理解哮喘的發病機理和治療的最新進展在amjrespircritcaremed.2008may15;177(10):1068-73中更全面地討論。
由于哮喘在兒童和成年人中如此普遍,因此一直需要能治療這種疾病或至少能緩解這種疾病所伴隨的癥狀而不會引起不需要的副作用的藥劑。同樣,一直需要用于治療copd的組合物和方法。本發明提供了用于治療copd和哮喘的組合物和方法以及詳細描述中所描述的其它優勢。
發明概述
本發明一方面提供了治療慢性阻塞性肺病的方法,包括給予copd患者治療有效量的v型膠原或其耐受原性片段。在本文所述方法的一個實施方案中,copd患者患有肺氣腫和/或慢性阻塞性支氣管炎。在本發明的另一實施方案中,v型膠原或其耐受原性片段通過口服給藥,并且可以以0.1mg-0.5mg的劑量給藥。在另一實施方案中,v型膠原或其耐受原性片段通過靜脈內給藥、通過肺內灌注給藥、通過吸入給藥或通過肌內給藥。在某些實施方案中,還可以使用不同途徑的組合。在本發明的另一實施方案中,所述方法還包括給予copd患者支氣管擴張藥、皮質類固醇或用于copd的其它已知治療。
本發明另一方面提供了治療哮喘的方法,包括給予哮喘患者治療有效量的v型膠原或其耐受原性片段。在所述方法的一個實施方案中,v型膠原或其耐受原性片段通過口服給藥,并且可以以0.1mg-0.5mg的劑量給藥。在其它實施方案中,v型膠原或其耐受原性片段通過靜脈內給藥、通過肺內灌注給藥、通過吸入給藥或通過肌內給藥。在另一實施方案中,所述方法還包括給予哮喘患者皮質類固醇、支氣管擴張藥和/或白三烯調節劑或用于哮喘的其它已知治療。
本發明另一方面提供了在有發展成慢性阻塞性肺病風險的個體中預防慢性阻塞性肺病的發展的方法,包括給予所述個體治療有效量的v型膠原或其耐受原性片段。在一個實施方案中,v型膠原或其耐受原性片段通過口服給藥,并且可以以0.1mg-0.5mg的劑量給藥。在其它實施方案中,v型膠原或其耐受原性片段通過靜脈內給藥、通過肺內灌注給藥、通過吸入給藥或通過肌內給藥,并且可以通過這些途徑的組合來給藥。
本發明另一方面提供了在有發展成哮喘風險的個體中預防哮喘的發展和惡化的方法,包括給予所述個體治療有效量的v型膠原或其耐受原性片段。在一個實施方案中,v型膠原或其耐受原性片段通過口服給藥,并且在某些實施方案中,可以以0.1mg-0.5mg的劑量給藥。在另一實施方案中,v型膠原或其耐受原性片段通過靜脈內給藥、通過肺內灌注給藥、通過吸入給藥、通過肌內給藥或通過這些途徑中一種或多種的組合來給藥。
本發明一方面提供了將copd或哮喘患者鑒定為用于v型膠原耐受療法的候選者的方法,包括將來自所述患者的血液樣品的至少一部分與v型膠原或其抗原性片段接觸;以及測量與v型膠原或其抗原性片段結合的抗體的水平(即測量v型膠原特異性抗體的水平);其中與v型膠原結合的抗體的存在指示copd或哮喘。在這方面中,v型膠原特異性抗體的水平可以與本文所述的其它臨床因素聯合用于診斷copd和哮喘。在一個實施方案中,將v型膠原或其抗原性片段結合于珠子上。在另一實施方案中,所述測量包括將與v型膠原或其抗原性片段結合的抗體與熒光標記的抗igg抗體接觸;以及通過流式細胞術檢測與結合于v型膠原上的抗體結合的熒光標記的抗igg抗體的量。
本發明另一方面提供了鑒定有發展成copd或哮喘風險的個體的方法,包括:將來自所述個體的血液樣品的至少一部分與v型膠原或其抗原性片段接觸;以及測量與v型膠原或其抗原性片段結合的抗體的水平(即測量v型膠原特異性抗體的水平);其中與v型膠原結合的抗體的存在與更高的風險相關,所述更高的風險比在沒有與v型膠原結合的抗體的個體中所預期的風險更高。在一個實施方案中,將v型膠原或其抗原性片段結合于珠子上。在另一實施方案中,所述測量包括將與v型膠原或其抗原性片段結合的抗體與熒光標記的抗igg抗體接觸;以及通過流式細胞術檢測與結合于v型膠原上的抗體結合的熒光標記的抗igg抗體的量。
在診斷或測量發展成copd或哮喘的風險的方法的某些實施方案中,所述方法中所用的抗igg抗體檢測所有igg亞型。在其它實施方案中,所述抗igg抗體特異性地檢測igg1亞型、或igg2亞型、或igg3亞型或igg4亞型。在這方面中,在疾病進展中可能發生一種亞型至另一種亞型的轉變,并且這可能指示疾病的惡化。因此,一種亞型隨時間增加可能指示疾病的惡化。
本發明另一方面提供了監測個體中copd或哮喘進展的方法,包括將來自所述個體的第一血液樣品的至少一部分與v型膠原或其抗原性片段接觸;測量第一血液樣品中與v型膠原或其抗原性片段結合的抗體的水平;將在較晚時間點采自所述個體的第二血液樣品的至少一部分與v型膠原或其抗原性片段接觸;測量第二血液樣品中與v型膠原或其抗原性片段結合的抗體的水平;以及將第二血液樣品中與v型膠原或其抗原性片段結合的抗體的水平與第一血液樣品中與v型膠原或其抗原性片段結合的抗體的水平進行比較;其中與第一樣品相比,第二樣品中與v型膠原結合的抗體的水平增加指示copd或哮喘在惡化,并且與第一樣品相比,第二樣品中與v型膠原結合的抗體的水平降低指示copd或哮喘的改善。copd和哮喘的其它臨床指征可以與本文所提供的方法聯合使用。在某些實施方案中,特定igg亞型(例如,igg1、igg2、igg3或igg4)的抗v型膠原抗體的增加指示copd或哮喘的進展。在某些實施方案中,將v型膠原或其抗原性片段結合于珠子上。在另一實施方案中,所述測量包括將與v型膠原或其抗原性片段結合的抗體與熒光標記的抗igg抗體接觸;以及通過流式細胞術檢測與結合于v型膠原上的抗體結合的熒光標記的抗igg抗體的量。在用于監測進展的方法的某些實施方案中,所述抗igg抗體檢測所有igg亞型。在其它實施方案中,所述抗igg抗體特異性地檢測igg1、igg2、igg3或igg4亞型。
結合以下詳細描述和附圖,本發明的這些方面和其它方面將變得明顯。
附圖簡要說明
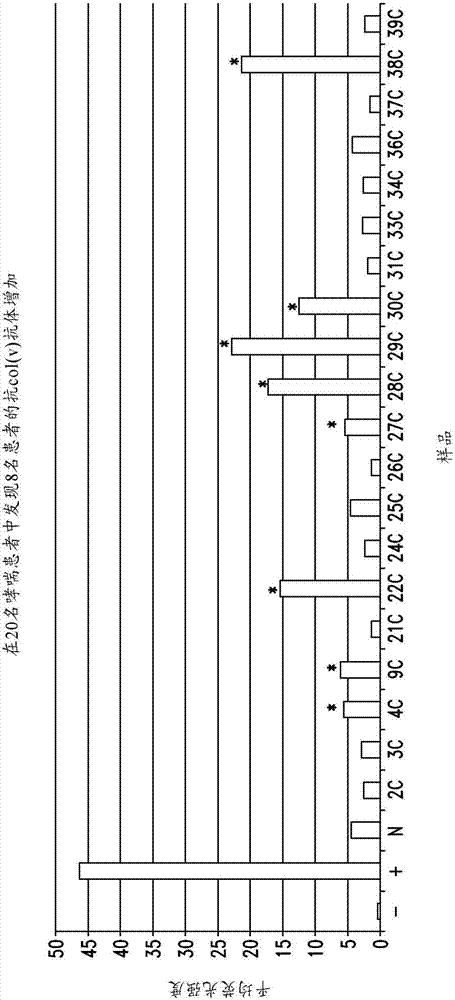
圖1是顯示copd患者中抗v型膠原抗體升高的柱形圖。
圖2是顯示哮喘患者中抗v型膠原抗體升高的柱形圖。
圖3顯示在靜脈內給予v型膠原之后,對小鼠中卵白蛋白誘導的氣道高反應的預防。除了colv(每個數據點n=2-5)之外所有組中的n=5。
圖4a-4e是顯示通過靜脈內給予col(v)而在肺單核細胞中誘導ifn-γ轉錄本的柱形圖。進行定量pcr來顯示il-4、il-5、il-13、ifn-γ和il-10細胞因子。只有col(v)iv在肺單核細胞中誘導ifn-γ轉錄本。數據表示從每組5只小鼠合并的肺單核細胞的rna。
發明的詳細描述
v型膠原
膠原蛋白是主要由羥脯氨酸(hyp)、甘氨酸(gly)和脯氨酸(pro)組成的重復氨基酸序列所形成的多肽鏈。膠原是人體中所見的最豐富的蛋白之一,在正常(非受傷)皮膚組織的真皮層中包含約80-85%的細胞外基質(ecm)。
基于序列同一性和功能,將膠原分成數種類型。i型、ii型和iii型膠原分子構成大多數動物細胞外結構的主要纖維。i型形成約90%的身體膠原,并且是骨、皮膚和腱的主要成分。ii型構成軟骨的主要纖維。膠原纖維以剛性板狀排列于骨中,以平行束狀排列于腱中,并以致密網狀排列于軟骨中。i型和較少量的iii型構成腱和皮膚。iv型膠原分子構成存在于基膜中的非常纖細、平滑的纖維。v型膠原(colv)是存在于肺中的小膠原(madriandfurthmayr,humanpathology,11:353-366,1980),并且位于細支氣管周的結締組織(madriandfurthmayr,am.j.pathol.,94:323-332,1979)、肺泡間質組織(konomietal.,1984)和毛細血管基膜(madriandfurthmayr,1979,同上)中。已知的其它膠原類型超過12種,但是還沒有得到很好的表征。
膠原多肽鏈的特征為由重復甘氨酸-x-y三聯體和球狀n-末端和c-末端結構域構成核心螺旋結構域。三條這樣的鏈以超螺旋彼此環繞,從而產生單個繩子樣的膠原分子。
以前的工作已經表明,對colv的自身免疫性與慢性同種移植異常(包括閉塞性細支氣管炎、閉塞性細支氣管炎綜合征(bos))、肺同種移植排斥和具有患ipf的風險相關(參見例如,美國專利7,348,005和wo2007/120947)。此外,這項工作表明colv的給予能誘導對同種抗原和colv的耐受(參見例如,wo2007/120947;圖10)。然而,在本發明之前,在哮喘或copd中沒有表明colv與自身免疫性的關聯。事實上,以前的發現表明,患copd的患者沒有抗col(v)的dth應答,這顯著不同于沒有患已知肺病的正常個體中所見的情況(參見wo2007/120947,實施例2;圖3)。因此,本發明是意料不到的,在于盡管以前觀察到患copd的患者與正常對照相比沒有升高的抗colv的dth應答,但是目前已經發現,copd患者具有升高的抗colv抗體(參見例如,實施例1、圖1)。
因此,本發明涉及在copd和哮喘患者中或有發展成這些疾病風險的個體中誘導對colv的耐受。
colv多核苷酸和多肽序列是本領域技術人員已知的,且可在公共數據庫獲得。本發明的示例性colv多核苷酸和多肽包括但不限于,智人膠原,v型,α1(col5a1),mrnancbi參照序列:nm_000093.3gi:89276750版本(seqidno:1);α1v型膠原前蛋白原[智人]:登錄號np_000084,gi:89276751版本(seqidno:2);智人膠原,v型,α2(col5a2),mrna,登錄號nm_000393,gi:89363016版本(seqidno:3);α2v型膠原前蛋白原[智人],登錄號np_000384,gi:89363017版本(seqidno:4);智人膠原,v型,α3(col5a3),mrna,登錄號nm_015719、nm_015719.3,gi:110735434(seqidno:5);膠原,v型,α3前蛋白原[智人],登錄號np_056534,np_056534.2版本,gi:110735435(seqidno:6)。
如本領域技術人員所認識到的,前原膠原在細胞中被加工成原膠原,該原膠原從細胞輸出并最終形成膠原原纖維和纖維。因此,本發明明確考慮原膠原和本文所述的已加工的或成熟形式的膠原蛋白。在這方面中,例如,seqidno:4的氨基酸1-26對應于信號肽,其在加工期間被切割,氨基酸27-1229是膠原α-2(v)鏈,且氨基酸1230-1499對應于c-末端前肽。本文具體公開的序列中的這些位置可被本領域技術人員識別,并且可通過其中提供序列批注的各種公共數據庫獲取。還應當注意,膠原蛋白的某些氨基酸在加工期間被修飾(例如,脯氨酸成為羥脯氨酸)。成熟的、修飾形式的v型膠原鏈,尤其是α-2鏈是本文明確考慮的。如其它地方所指出的,本發明中所用的v型膠原及其α鏈可以從多種來源純化或重組產生。
本文所用的術語“多肽”以其常規含義使用,即作為氨基酸的序列。多肽并不限制于特定長度的產物,因此,多肽定義包括肽、寡肽和蛋白,并且這類術語本文可互換使用,除非明確指出不是這樣。該術語還不指或排除多肽的表達后修飾,例如,天然存在的或非天然存在的糖基化、乙酰化、磷酸化等以及領域內已知的其它修飾。多肽可以是整個蛋白或其子序列。本發明背景下的特定目的多肽是包含耐受原性片段的氨基酸子序列。
另一方面,本發明提供了這樣的多肽片段,該多肽片段包含本文所示的膠原多肽組合物的至少約5、10、15、20、25、50或100個連續氨基酸或更多,包括所有中間長度,所述膠原多肽組合物例如seqidno:2、4或6所示的那些或seqidno:1、3或5序列中所示的多核苷酸序列所編碼的那些。
另一方面,本發明提供了本文所述的多肽組合物的變體。本發明通常包括的多肽變體沿其長度與本文所示的多肽序列通常表現出至少約70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%或更高的同一性(按下述確定)。
在一個實施方案中,本發明提供的多肽片段和變體是本文所述的免疫學上耐受原性的。
本文所用的術語多肽“變體”是通過一個或多個取代、缺失、添加和/或插入而通常不同于本文具體公開的多肽的多肽。這類變體可以是天然存在的或可以是合成產生的,例如通過修飾一個或多個本發明的上述多肽序列,并按本文所述評價它們的耐受原活性和/或利用領域內公知的任何技術。
在很多情況下,變體含有保守型取代。“保守型取代”是指氨基酸被具有相似性質的另一個氨基酸所取代,使得肽化學領域內的技術人員可預料到該多肽的二級結構和親水性基本未改變。可以在本發明的多核苷酸和多肽結構中進行修飾,并仍能獲得編碼具有期望特性(例如,具有耐受原性特性)的變體或衍生多肽的功能性分子。當希望改變多肽的氨基酸序列來產生等同的甚至改進的本發明多肽的耐受原性變體或部分時,本領域技術人員通常按照表1改變編碼dna序列的一個或多個密碼子。
例如,在蛋白結構中,某些氨基酸可以被其他的氨基酸取代,而不會造成與諸如抗體的抗原結合區或基底分子的結合位點的結構的相互結合能力的明顯損失。由于是蛋白的相互作用能力和性質限定了該蛋白的生物學功能活性,因此可以在蛋白序列中進行某些氨基酸序列的取代,以及當然也可在其基礎dna編碼序列中進行取代,并仍可獲得具有相似性質的蛋白。因而已考慮到,可以在所公開的組合物的肽序列或編碼所述肽的相應dna序列中進行各種改變,而不會造成它們的耐受原性功效或活性的明顯損失。
表1
在進行這些改變時,應當考慮氨基酸的親水指數。本領域內普遍理解親水氨基酸指數在賦予蛋白相互作用的生物學功能中的重要性(kyteanddoolittle,1982,通過引用并入本文)。已接受氨基酸的相對親水特性與得到的蛋白的二級結構有關,而該二級結構隨之限定了該蛋白與諸如酶、底物、受體、dna、抗體、抗原等其它分子的相互作用。基于氨基酸的疏水性和電荷特性,每種氨基酸具有指定的親水指數(kyteanddoolittle,1982)。這些值是:異亮氨酸(+4.5)、纈氨酸(+4.2)、亮氨酸(+3.8)、苯丙氨酸(+2.8)、半胱氨酸/胱氨酸(+2.5)、蛋氨酸(+1.9)、丙氨酸(+1.8)、甘氨酸(-0.4)、蘇氨酸(-0.7)、絲氨酸(-0.8)、色氨酸(-0.9)、酪氨酸(-1.3)、脯氨酸(-1.6)、組氨酸(-3.2)、谷氨酸(-3.5)、谷氨酰胺(-3.5)、天冬氨酸(-3.5)、天冬酰胺(-3.5)、賴氨酸(-3.9)和精氨酸(-4.5)。
本領域內已知,某些氨基酸可以被具有相似親水指數或評分的其它氨基酸取代,而仍產生具有相似生物學活性的蛋白,即仍獲得生物學功能等同的蛋白。在進行這些改變時,親水指數在±2以內的氨基酸的取代是優選的,特別優選的是在±1以內的氨基酸的取代,并且更特別優選的是在±0.5以內的那些氨基酸的取代。美國專利4,554,101(具體通過引用全文并入本文)陳述,蛋白最局部的平均親水性(由其鄰近氨基酸的親水性導致)與該蛋白的生物學性質相關。
如美國專利第4,554,101號詳述,已將下述親水性值指定給氨基酸殘基:精氨酸(+3.0)、賴氨酸(+3.0)、天冬氨酸(+3.0±1)、谷氨酸(+3.0±1)、絲氨酸(+0.3)、天冬酰胺(+0.2)、谷氨酰胺(+0.2)、甘氨酸(0)、蘇氨酸(-0.4)、脯氨酸(-0.5±1)、丙氨酸(-0.5)、組氨酸(-0.5)、半胱氨酸(-1.0)、蛋氨酸(-1.3)、纈氨酸(-1.5)、亮氨酸(-1.8)、異亮氨酸(-1.8)、酪氨酸(-2.3)、苯丙氨酸(-2.5)、色氨酸(-3.4)。應當理解,氨基酸可以被具有相似親水性值的另一氨基酸取代,且仍獲得生物學上等同的,尤其是免疫學上等同的蛋白。在這類改變中,親水性值在±2以內的氨基酸的取代是優選的,特別優選的是在±1以內的氨基酸的取代,并且更特別優選的是在±0.5以內的那些氨基酸的取代。
如上文所述,氨基酸取代因而通常基于氨基酸側鏈取代基的相對的相似性,例如,它們的疏水性、親水性、電荷、大小等。綜合考慮了上述各種特性的示例性取代是本領域技術人員所公知的,并且包括:精氨酸與賴氨酸、谷氨酸與天冬氨酸、絲氨酸與蘇氨酸、谷氨酰胺與天冬酰胺、以及纈氨酸、亮氨酸與異亮氨酸。
此外,可以進一步修飾任何多核苷酸以增加體內穩定性。可能的修飾包括但不限于,在5'和/或3'末端添加側翼序列、在主鏈中使用硫代磷酸或2'o-甲基而不是磷酸二酯酶連接和/或包含諸如肌苷、q核苷和和懷丁苷的非常規堿基以及乙酰基、甲基、巰基和其它修飾形式的腺嘌呤、把嘧啶、鳥嘌呤、胸腺嘧啶和尿嘧啶。
還可以基于殘基的極性、電荷、溶解性、疏水性、親水性和/或兩性性質的相似性進行氨基酸取代。例如,帶負電荷的氨基酸包括天冬氨酸和谷氨酸,帶正電荷的氨基酸包括賴氨酸和精氨酸,以及帶有具有相似親水性值的不帶電荷極性頭部基團的氨基酸包括:亮氨酸、異亮氨酸和纈氨酸,甘氨酸和丙氨酸,天冬酰胺和谷氨酰胺,以及絲氨酸、蘇氨酸、苯丙氨酸和酪氨酸。可以代表保守型改變的其他氨基酸組包括:(1)ala、pro、gly、glu、asp、gln、asn、ser、thr;(2)cys、ser、tyr、thr;(3)val、ile、leu、met、ala、phe;(4)lys、arg、his;和(5)phe、tyr、trp、his。變體還可以含有非保守型改變或可選擇地含有非保守型改變。在優選的實施方案中,變體多肽與天然序列的區別在于5個或更少的氨基酸的取代、缺失或添加。還可以(或可選擇地)通過例如對多肽的耐受原性、二級結構和親水性具有最小影響的氨基酸的缺失或添加來修飾變體。
如上文所述,多肽在蛋白的n末端可以包含信號(或前導)序列,其在翻譯的同時或在翻譯后指導蛋白的轉移。多肽還可以與連接物或其他序列綴合,以便于多肽的合成、純化或鑒定(例如,多聚組氨酸),或用于增強多肽與固相載體的結合。例如,多肽可以與免疫球蛋白fc區綴合。
當比較多肽序列時,如果兩個序列中的氨基酸序列在如下文所述進行最大對應比對時是相同的,則可以認為所述兩個序列是“同一的”。通常通過在比較窗中比較序列來進行兩個序列間的比較,從而鑒定和比較序列相似性的局部區域。本文所用的“比較窗”是指至少約20個連續位置、通常30-約75個、40-約50個連續位置的節段,其中在將序列與具有相同數量連續位置的參考序列最佳比對后,可以將所述序列與參考序列進行比較。
可以使用lasergene生物信息學軟件套裝中的megalign程序(dnastar,inc.,madison,wi),使用默認參數,進行用于比較的序列的最佳比對。該程序包括以下參考文獻中描述的數種比對方案:dayhoff,m.o.(1978)amodelofevolutionarychangeinproteins-matricesfordetectingdistantrelationships(蛋白質的進化改變模型-用于檢驗遠距離關系的矩陣);indayhoff,m.o.(ed.)atlasof蛋白sequenceandstructure(蛋白質序列和結構圖譜),nationalbiomedicalresearchfoundation,washingtondcvol.5,suppl.3,pp.345-358;heinj.(1990)unifiedapproachtoalignmentandphylogenes(比對和系統發生的標準方法)pp.626-645;methodsinenzymology(酶學方法)vol.183,academicpress,inc.,sandiego,ca;higgins,d.g.andsharp,p.m.,cabios5:151-153(1989);myers,e.w.andmullerw.,cabios4:11-17(1988);robinson,e.d.,comb.theor11:105(1971);saitou,n.nei,m.,mol.biol.evol.4:406-425(1987);sneath,p.h.a.andsokal,r.r.,numericaltaxonomy–theprinciplesandpracticeofnumericaltaxonomy(數值分類法-數值分類法的原理和實踐),freemanpress,sanfrancisco,ca(1973);wilbur,w.j.andlipman,d.j.,proc.natl.acad.,sci.usa80:726-730(1983)。
可選擇地,可以通過以下來進行用于比較的序列的最佳比對:smithandwatermanadd.apl.math2:482(1981)的局部同一性算法、needlemanandwunschj.mol.biol.48:443(1970)的同一性比對算法、pearsonandlipmanproc.natlacad.sci.usa85:2444(1988)的相似性方法搜索、這些算法的計算機執行(wisconsin遺傳學軟件包中的gap、bestfit、blast、fasta和tfasta,geneticscomputergroup(gcg),575sciencedr.,madison,wi)、或檢查。
適于確定序列同一性和序列相似性百分比的算法的一個優選實例是blast和blast2.0算法,其分別描述于altschuletal.nucl.acidsres.25:3389-3402(1977)和altschuletal.j.mol.biol.215:403-410(1990)中。例如,可以按照本文所述的參數使用blast和blast2.0,從而確定本發明多核苷酸和多肽的序列同一性百分比。通過美國國家生物技術信息中心(nationalcenterforbiotechnologyinformation)可以公開獲得進行blast分析的軟件。對于氨基酸序列而言,可以將評分矩陣用于計算累積評分。在以下情況時停止每個方向的字符命中延伸:當累積比對評分從其最大達到的值下降量x時;由于一個或多個負評分殘基比對的累積而使累積評分達到0或低于0時;或達到任一序列的末端時。blast算法參數w、t和x決定了所述比對的靈敏度和速度。
在一個優選的方法中,通過在至少20個位置的比較窗中比較兩個最佳比對的序列來確定“序列同一性百分比”,其中,與用于兩個序列最佳比對的參考序列(其不包含添加或缺失)相比,比較窗中的多肽序列的一部分可以包含20%或更少的、通常為5%-15%、或10%-12%的添加或缺失(即,空位)。通過以下來計算百分比:確定兩個序列中都存在的相同氨基酸殘基的位置數量來產生配對位置數,將配對位置數除以參考序列中(即窗的大小)的位置總數,并且將結果乘以100以產生序列同一性的百分比。
在其它示例性的實施方案中,多肽可以是包含多個本文所述的多肽的融合多肽,或者可以是包含至少一個本文所述的多肽和一段無關序列(例如用于純化的組氨酸標簽或導向肽)的融合多肽。例如,融合伴侶可以有助于以高于天然重組蛋白的產量表達蛋白(表達增強子)。可以選擇其它的融合伴侶,以增加多肽的溶解性或使多肽能靶向于所需的細胞內區域。其它融合伴侶包括促進多肽純化的親和性標簽。在某些實施方案中,融合伴侶能增加多肽的耐受原性或提高其通過細胞的攝入。在其它實施方案中,融合伴侶包含免疫應答增強子。
通常可以用標準技術制備融合蛋白,包括化學綴合。優選地,在表達系統中,融合多肽作為重組多肽表達,允許以高于非融合多肽的水平產生。簡單而言,可以分別組裝編碼多肽組分的dna序列,并將其連入合適的表達載體中。使用或不使用肽連接物將編碼一種多肽組分的dna序列的3'端與編碼第二多肽組分的dna序列的5'端連接,這樣序列的讀碼框是同相的。這允許翻譯為同時保留兩個組分多肽的生物活性的單一融合多肽。
可以用肽連接物序列將第一和第二多肽組分隔開足以確保每種多肽折疊成其二級和三級結構的距離。用本領域內公知的標準技術將這類肽連接物序列并入融合多肽。可以基于下列因素選擇合適的肽連接物序列:(1)它們采取柔韌的、伸展的構象的能力;(2)它們不會采取與第一和第二多肽上的功能表位相互作用的二級結構的能力;和(3)可以與所述多肽功能表位反應的疏水或帶電殘基的缺乏。優選的肽連接物序列包含gly、asn和ser殘基。諸如thr和ala的其它接近中性的氨基酸也可以用于連接物序列中。可以有效地用作連接物的氨基酸序列包括公開于marateaetal.,gene40:39-461985;murphyetal.,proc.natl.acad.sci.usa83:8258-82621986;美國專利第4,935,233號和第4,751,180號中的那些氨基酸序列。通常,連接物序列的長度可以為1-約50個氨基酸。當第一和第二多肽具有可以用來隔開功能結構域和防止空間干擾的非必需n末端氨基酸區時,不需要連接物序列。
將連接的dna序列可操縱地連接于合適的轉錄或翻譯調節元件。負責dna表達的調節元件僅位于編碼第一多肽的dna序列的5'。同樣,終止翻譯所需的終止密碼子和轉錄終止信號僅位于編碼第二多肽的dna序列的3'。
本發明的一個實施方案涉及融合多肽和編碼它們的多核苷酸。其中所述融合伴侶包含能引導多肽進入內體/溶酶體區域的導向信號,如美國專利第5,633,234號中所描述。當與該導向信號融合時,本發明的耐受原性多肽會更有效地聯合mhcii類分子,從而為該多肽特異的合適的cd4+t細胞提供增強的體內刺激。
可以利用各種公知的合成和/或重組技術制備本發明的多肽,后者將在下文進一步描述。利用本領域技術人員公知的技術通過合成方法產生的多肽、一部分或其它變體通常小于約150個氨基酸。在一個示例性的實例中,利用任何商業上可獲得的固相技術來合成這類多肽,例如merrifield固相合成方法,其中氨基酸被相繼添加到生長的氨基酸鏈上。參見merrifield,j.am.chem.soc.85:2149-2146,1963。用于多肽自動合成的設備可購自諸如perkinelmer/appliedbiosystemsdivision(fostercity,ca)的供應商,并且可以按照廠商的說明書進行操縱。
通常,本發明的多肽組合物(包括融合多肽)是分離的。“分離的”多肽是從其原來的環境中移出的多肽。例如,如果將天然存在的蛋白或多肽與天然系統中的部分或所有的共存物質分開,則該天然存在的蛋白或多肽是分離的。優選地,這類多肽還被純化,例如是至少約90%純的,更優選為至少約95%純的和最優選為至少約99%純的。
colv蛋白可以從多種來源純化或可以從商業來源(collaborativebiomedicalproducts/becton,dickinsonandcompany,franklinlakes,njusa)購買。為了實施一些實施方案,可能必須獲得純的或基本純的膠原或其耐受原性片段、表位或抗原部分。通過多種途徑可易于獲取這些材料例如v型膠原或其片段,包括但不限于動物來源、人尸體或重組方式。其它方法包括部分消化諸如v型膠原的膠原。在這方面中,人類v型膠原可從人尸體或其它來源提取,并通過差別nacl沉淀來純化(seyerandkang,1989)。例如,將胎盤組織切碎、洗滌并懸浮于含有0.2mnacl的0.5m乙酸中,并于4℃用胃蛋白酶消化。從離心的樣本吸出上清,收集沉淀并重復提取方案。將兩次消化的上清混合,并通過差別nacl沉淀從0.5m乙酸上清中純化col(v)(smithetal.,1985;seyerandkang,1989)。v型膠原通常可溶于0.7mnacl中,并在1.2mnacl中沉淀。
對于這些需要純化α(v)鏈的實施方案而言,可以重復乙酸溶解和nacl沉淀的循環,直到獲得具有α-鏈比值、α1(v)/α2(v)大約為2的v型制劑,該比值是通過sds-聚丙烯酰胺凝膠電泳(smithetal.,1985)或本領域技術人員已知的其它合適的方法來測定的。可以通過deae-纖維素層析(seyerandkang,1989)或本領域技術人員已知的其它方法,例如proteinpurificationprotocols,ed.shawndoonan,humanapress,1996)中所描述的那些方法實現α1(v)與α2(v)的分離。可從柱洗提α1(v)與α2(v)鏈,并通過之前所報道的sds-聚丙烯酰胺凝膠電泳(smith,jr.etal,1985)來確認純度。可以將完整的col(v)或α1(v)與α2(v)鏈稀釋于pbs(0.5mg/ml)或其它合適的緩沖液中直到使用。
在某些實施方案中,本發明提供了編碼本發明膠原蛋白的多核苷酸。示例性的多核苷酸是seqidno:1、3和5中所示的那些和它們編碼本文所述的膠原蛋白的耐受原性片段的片段。
術語“dna”和“多核苷酸”在本文實質上是可交換使用的,是指已從特定物種的總基因組dna分離出的dna分子。本文所用的“分離的”表示多核苷酸基本上遠離其它編碼序列,以及該dna分子不含大部分無關的編碼dna,例如大量的染色體片段或其它功能基因或多肽編碼區。當然,這是指最初分離的dna分子,并且不排除后來通過人工添加到該節段上的基因或編碼區。
如本領域技術人員所理解的,本發明的多核苷酸組合物包括基因組序列、基因組外序列和質粒編碼的序列以及表達或被改造來表達蛋白、多肽、肽等的較小的工程基因節段。這類區段可以是天然分離的,或人工合成修飾的。
可以利用任何成熟的技術來鑒定、制備和/或操縱本發明的多核苷酸組合物(通常參見,sambrooketal.,molecularcloning:alaboratorymanual(分子克隆:實驗手冊),coldspringharborlaboratories,coldspringharbor,ny,1989;ausubeletal.(2001currentprotocolsinmolecularbiology(現代分子生物學技術),greenepubl.assoc.inc.&johnwiley&sons,inc.,ny,ny)及其它類似的參考文獻)。
可利用多種模板依賴性的方法來擴增樣品中所存在目的靶序列。公知的擴增方法之一是聚合酶鏈式反應(pcrtm),這在美國專利第4,683,195號、第4,683,202號和第4,800,159號中有詳細描述,通過引用將每一篇全文并入本文。
任何其它模板依賴性的方法,其中多數是pcrtm擴增技術的衍化是領域內已知且易于利用的。舉例來說,一些這類方法包括例如描述于歐洲專利申請公開號320,308和美國專利第4,883,750號中的連接酶鏈式反應(稱為lcr)、描述于pct國際專利申請公開號pct/us87/00880中的qβ復制酶、鏈置換擴增(sda)和修復鏈反應(rcr)。其它擴增方法描述于英國專利申請號2202328和pct國際專利申請公開號pct/us89/01025中。其它核酸擴增方法包括基于轉錄的擴增系統(tas)(pct國際專利申請公開號wo88/10315),包括基于核酸序列的擴增(nasba)和3sr。歐洲專利申請公開號329,822描述了涉及循環合成單鏈rna(“ssrna”)、ssdna和雙鏈dna(dsdna)的核酸擴增方法。pct國際專利申請公開號wo89/06700描述了基于啟動子/引物序列與靶單鏈dna(“ssdna”)雜交,隨后轉錄出該序列的很多rna拷貝的核酸序列擴增方案。諸如“race”(frohman,1990)和“單側pcr”(ohara,1989)的其它擴增方法也是本領域技術人員公知的。
利用公知技術可以使用本發明多核苷酸的擴增部分從合適的文庫(例如,腫瘤cdna文庫)分離出全長基因。
還如本領域技術人員所公認的,本發明的多核苷酸可以是單鏈的(編碼或反義)或雙鏈的,并且可以是dna(基因組、cdna或合成物)或rna分子。rna分子可以包括hnrna分子和mrna分子,所述hnrna分子包含內含子并且以一對一的方式對應于dna分子,而所述mrna分子不含內含子。其它編碼或非編碼序列可以,但不需要,存在于本發明的多核苷酸中,并且多核苷酸可以,但不需要,與其他的分子和/或支持物質連接。
多核苷酸可以包含天然序列(即,編碼本發明的多肽/蛋白或其一部分的內源序列),或可以包含編碼這類序列的變體或衍生物,優選耐受原性變體或衍生物的序列。
在其它相關的實施方案中,本發明提供了與本文在seqidno:1、3和5中所提供的序列具有大量同一性的多核苷酸變體,例如,利用本文所述的方法(例如,如下文討論的,使用標準參數的blast分析),包含與本發明的多核苷酸序列相比具有至少70%的序列同一性,優選至少75%、80%、85%、90%、95%、96%、97%、98%或99%或更高的序列同一性的那些序列。本領域技術人員應當認識到,通過考慮密碼簡并性、氨基酸相似性、讀碼框定位等,這些值可以被適當調整以確定兩個核苷酸序列所編碼的蛋白的對應的同一性。
通常,多核苷酸變體可以包含一個或多個取代、添加、缺失和/或插入,優選使得由該變體多核苷酸所編碼的多肽的耐受原活性相對于本文具體展示的多核苷酸序列所編碼的多肽沒有顯著減弱。術語“變體”還應當被理解為包括異種來源的同源基因。
在其它實施方案中,本發明提供了多核苷酸片段,其包含與本文所公開的一個或多個序列相同或互補的序列的不同長度的連續區段,或由這些區段組成。例如,本發明提供的多核苷酸包含至少約10、15、20、30、40、50、75、100、150、200、300、400、500或1000或更多個本發明所公開的一個或多個序列的連續核苷酸,以及所有的中間長度,或者由至少約10、15、20、30、40、50、75、100、150、200、300、400、500或1000或更多個本發明所公開的一個或多個序列的連續核苷酸,以及所有的中間長度的連續核苷酸組成。應當容易理解,此處的“中間長度”表示所提到的值之間的任何長度,例如16、17、18、19等;21、22、23等;30、31、32等;50、51、52、53等;100、101、102、103等;150、151、152、153等;包括跨越200-500、500-1,000等的所有整數。可以在本文所述的多核苷酸序列的一端或兩端延伸天然序列中未見的其它核苷酸。位于所公開序列的任一端或位于所公開序列的兩端的這個其它序列可以由1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20個核苷酸組成。
在本發明的另一實施方案中,提供能夠在中度至高度嚴緊條件下與本文提供的多核苷酸序列,或該多核苷酸的片段,或該多核苷酸的互補序列雜交的多核苷酸組合物。雜交技術是分子生物學領域內公知的。為了說明的目的,適于測試本發明的多核苷酸與其他多核苷酸雜交的中度嚴緊條件包括:在5×ssc、0.5%sds、1.0mmedta(ph8.0)的溶液中預先洗滌;在5×ssc中、于50℃-60℃過夜雜交;然后,用含有0.1%sds的2×ssc、0.5×ssc和0.2×ssc中的每一種、于65℃洗滌20分鐘,洗滌兩次。本領域技術人員應當理解,容易操縱雜交的嚴緊度,例如通過改變雜交溶液的鹽含量和/或改變進行雜交時的溫度。例如,在另一實施方案中,合適的高度嚴緊雜交條件包括上文所討論的那些條件,除此之外,要將雜交溫度提高到例如60-65℃或65-70℃。
在一個實施方案中,這類多核苷酸變體編碼這樣的多肽,其具有本文具體展示的多肽序列的至少約50%、優選至少約70%,且更優選至少約90%、95%、96%、97%、98%、99%或更高的耐受原活性水平。
不考慮編碼序列自身的長度,本發明的多核苷酸或其片段可以與其它dna序列組合,所述其它dna序列例如啟動子、聚腺苷酸化信號、其它限制性酶切位點、多克隆位點、其它編碼節段等,從而它們的總長度可以有相當大的變化。因此,應當考慮到,可以使用幾乎任何長度的核酸片段,總長度優選地受在計劃的重組dna流程中制備和使用的便利性的限制。例如,本發明的多種實施中可考慮使用總長度為約10,000、約5000、約3000、約2,000、約1,000、約500、約200、約100、約50個堿基對等(包括所有中間長度)的示例性的多核苷酸節段。
當比較多核苷酸序列時,如果兩個序列中的核苷酸序列在如下文所述進行最大對應比對時是相同的,則可以認為所述兩個序列是“同一的”。通常通過在比較窗中比較序列來進行兩個序列間的比較,從而鑒定和比較序列相似性的局部區域。本文所用的“比較窗”是指至少約20個連續位置、通常30-約75個、40-約50個連續位置的節段,其中在將序列與具有相同數量連續位置的參考序列最佳比對后,可以將所述序列與參考序列進行比較。
可以使用lasergene生物信息學軟件套裝中的megalign程序(dnastar,inc.,madison,wi),使用默認參數,進行用于比較的序列的最佳比對。該程序包括以下參考文獻中描述的數種比對方案:dayhoff,m.o.(1978)amodelofevolutionarychangeinproteins-matricesfordetectingdistantrelationships(蛋白質的進化改變模型-用于檢驗遠距離關系的矩陣);indayhoff,m.o.(ed.)atlasof蛋白sequenceandstructure(蛋白質序列和結構圖譜),nationalbiomedicalresearchfoundation,washingtondcvol.5,suppl.3,pp.345-358;heinj.unifiedapproachtoalignmentandphylogenes(比對和系統發生的標準方法)pp.626-645(1990);methodsinenzymology(酶學方法)vol.183,academicpress,inc.,sandiego,ca;higgins,d.g.andsharp,p.m.,cabios5:151-153(1989);myers,e.w.andmullerw.,cabios4:11-17(1988);robinson,e.d.,comb.theor11:105(1971);saitou,n.nei,m.,mol.biol.evol.4:406-425(1987);sneath,p.h.a.andsokal,r.r.,numericaltaxonomy–theprinciplesandpracticeofnumericaltaxonomy(數值分類法-數值分類法的原理和實踐),freemanpress,sanfrancisco,ca(1973);wilbur,w.j.andlipman,d.j.,proc.natl.acad.,sci.usa80:726-730(1983)。
可選擇地,可以通過以下來進行用于比較的序列的最佳比對:smithandwatermanadd.apl.math2:482(1981)的局部同一性算法、needlemanandwunschj.mol.biol.48:443(1970)的同一性比對算法、pearsonandlipmanproc.natlacad.sci.usa85:2444(1988)的相似性方法搜索、這些算法的計算機執行(wisconsin遺傳學軟件包中的gap、bestfit、blast、fasta和tfasta,geneticscomputergroup(gcg),575sciencedr.,madison,wi)、或檢查。
適于確定序列同一性和序列相似性百分比的算法的一個優選實例是blast和blast2.0算法,其分別描述于altschuletal.nucl.acidsres.25:3389-3402(1977)和altschuletal.j.mol.biol.215:403-410(1990)中。例如,可以按照本文所述的參數使用blast和blast2.0,從而確定本發明多核苷酸的序列同一性百分比。通過美國國家生物技術信息中心(nationalcenterforbiotechnologyinformation)可以公開獲得進行blast分析的軟件。在一示例性的實例中,對于核苷酸序列而言,可以使用參數m(對于一對配對殘基的獎勵分;總是>0)和n(對于不配對殘基的罰分;總是<0)來計算累積評分。在以下情況時停止每個方向的字符命中延伸:當累積比對評分從其最大達到的值下降量x時;由于一個或多個負評分殘基比對的累積而使累積評分達到0或低于0時;或達到任一序列的末端時。blast算法參數w、t和x決定了所述比對的靈敏度和速度。blast程序(對于核苷酸序列而言)使用以下默認值:字長(w)11、期望(e)10、blosum62評分矩陣(參見henikoffandhenikoff,proc.natl.acad.sci.usa89:10915(1989))比對、(b)50、期望(e)10、m=5、n=-4和雙鏈比較。
優選地,通過在至少20個位置的比較窗中比較兩個最佳比對的序列來確定“序列同一性百分比”,其中,與用于兩個序列最佳比對的參考序列(其不包含添加或缺失)相比,比較窗中的多核苷酸序列的一部分可以包含20%或更少的、通常為5%-15%、或10%-12%的添加或缺失(即,空位)。通過以下來計算百分比:確定兩個序列中都存在的相同核酸堿基的位置數量來產生配對位置數,將配對位置數除以參考序列中(即窗的大小)的位置總數,并且將結果乘以100以產生序列同一性的百分比。
本領域技術人員應當理解,由于遺傳密碼的簡并性,有很多核苷酸序列編碼本文所述的多肽。這些多核苷酸中的某些與任何天然基因的核苷酸序列具有低的同源性。雖然如此,本發明特別考慮到由于密碼子使用中的差異而不同的多核苷酸。此外,包含本文提供的多核苷酸序列的基因的等位基因包括在本發明范圍內。等位基因是由于一個或多個諸如核苷酸的缺失、添加和/或取代的突變而改變的內源基因。所得到的mrna和蛋白可以,但不需要,具有改變的結構或功能。可以用標準技術鑒定等位基因(例如雜交、擴增和/或數據庫序列比較)。
因此,在本發明的另一實施方案中,使用諸如定點誘變的誘變方法來制備本文所述的多肽的耐受原性變體和/或衍生物。通過該方法,可以通過對編碼多肽序列的基礎多核苷酸進行誘變而在該多肽序列中產生特定的修飾。這些技術為制備并測試序列變體提供了簡易的方法,例如通過在多核苷酸中導入一個或多個核苷酸序列改變而并入一個或多個上述的修飾。
定點誘變允許通過使用能編碼所需突變的dna序列的特定寡核苷酸序列以及足夠數量的鄰近核苷酸來產生突變體,所述鄰近核苷酸用于提供大小和序列復雜性足以在待跨越的缺失位點的兩側形成穩定雙螺旋的引物序列。可以在所選的多核苷酸序列中利用突變來提高、改變、減低、修飾或以其它方式改變該多核苷酸自身的性質和/或改變所編碼的多肽的性質、活性、組成、穩定性或一級序列。
在本發明的某些實施方案中,本發明人考慮對所公開的多核苷酸序列進行誘變,從而改變所編碼多肽的一種或多種性質,例如多肽的耐受原性。定點誘變技術是領域內公知的,且廣泛用于產生多肽和多核苷酸兩者的變體。例如,定點誘變常用來改變dna分子的特定部分。在這類實施方案中,利用通常包含約14至約25個核苷酸或大約長度的引物,在待改變的序列位點的兩側均具有約5至約10個殘基。
如本領域技術人員所理解的,定點誘變技術通常使用既能以單鏈又能以雙鏈形式存在的噬菌體載體。用于定點誘變中的典型載體包括諸如m13噬菌體的載體。這些噬菌體容易買到,并且其使用通常是本領域技術人員公知的。雙鏈質粒也常用于定點誘變中,其排除了將目的基因從質粒轉入噬菌體中的步驟。
通常,首先通過獲得單鏈載體或雙鏈載體的解鏈的兩條鏈來進行本文的定點誘變,所述載體在其序列中包含編碼所需肽的dna序列。通常通過合成法制備攜帶所需突變序列的寡核苷酸引物。然后將該引物與單鏈載體退火,并利用諸如大腸桿菌聚合酶iklenow片段的酶進行dna聚合,從而完成突變攜帶鏈的合成。因此,形成其中一條鏈編碼原始非突變序列和第二鏈攜帶所需突變的異源雙螺旋。然后,用該該異源雙螺旋載體轉化合適的細胞,例如大腸桿菌細胞,并選擇包含攜帶突變序列排列的重組載體的克隆。
利用定點誘變制備編碼所選肽的序列變體的dna節段提供了一種產生具有潛在用途的種類的工具,并且這并不意味著是受限的,因為還有可以獲得肽的序列變體和編碼該變體的dna序列的其它方式。例如,可以用諸如羥胺的誘變劑處理編碼所需肽序列的重組載體,從而獲得序列變體。關于這些方法和方案的具體詳述見于maloyetal.,1994;segal,1976;prokopandbajpai,1991;kuby,1994;和maniatisetal.,1982的教導,將每一篇通過引用并入本文,用于該目的。
本文所用的術語“寡核苷酸定點誘變方法”是指模板依賴的過程和載體介導的增殖,該增殖導致特定核酸分子的濃度相對于其初始濃度有所提高,或導致可檢測信號濃度的提高,例如擴增。本文所用的術語“寡核苷酸定點誘變方法”意指涉及引物分子的模板依賴性延伸的過程。術語模板依賴的過程是指rna或dna分子的核酸合成,其中新合成的核酸鏈的序列由公知的互補性堿基配對原則決定(參見例如,watson,1987)。通常,載體介導的方法涉及將核酸片段導入dna或rna載體中、載體的克隆擴增和擴增核酸片段的回收。美國專利第4,237,224號提供了這類方法的實例,通過引用將其全文并入本文。
在用于產生本發明多肽變體的另一方法中,可以利用如美國專利第5,837,458號所述的循環序列重組。在該方法中,進行重組和篩選或選擇的重復循環,從而“演化出”本發明的具有例如增強的耐受原活性的個體多核苷酸變體。
在本發明的其它實施方案中,編碼本發明多肽或其融合蛋白或功能等同物的多核苷酸序列或多核苷酸序列片段可以用于重組dna分子中,從而指導多肽在合適的宿主細胞中的表達。由于遺傳密碼內在的簡并性,可以產生編碼基本上相同的或功能上等同氨基酸序列的其它dna序列,并且這些序列可以用來克隆和表達給定的多肽。
如本領域技術人員所理解地,在一些情況下,產生具有非天然存在的密碼子的編碼多肽的核苷酸序列是有利的。例如,可以選擇特定原核生物或真核生物宿主所偏好的密碼子來增加蛋白表達率或產生具有所需特性的重組rna轉錄本,所述所需特性例如比天然存在的序列所產生的轉錄本更長的半衰期。
此外,可以使用本領域內公知的方法改造本發明的多核苷酸序列,從而為了多種原因改變多肽編碼序列,包括但不限于修飾基因產物的克隆、加工和/或表達的改變。例如,通過隨機斷裂和基因片段和合成寡核苷酸的pcr組裝而進行的dna改組可用來改造核苷酸序列。此外,定點誘變可用來插入新的限制性位點、改變糖基化模式、改變密碼子偏好性、產生剪接變體或導入突變等。
在本發明的另一實施方案中,天然的、修飾的或重組的核酸序列可連接至異源序列,從而編碼融合蛋白。例如,為了篩查作為多肽活性抑制劑的肽文庫,編碼能用商購抗體識別的嵌合蛋白可能是有用的。還可以對融合蛋白進行改造使之在多肽編碼序列和異源蛋白序列之間含有切割位點,以便該多肽可被切割并從所述異源部分純化出來。
可以利用領域內公知的化學方法全部或部分合成編碼所需多肽的序列(參見caruthers,m.h.etal.(1980)nucl.acidsres.symp.ser.215-223,horn,t.etal.(1980)nucl.acidsres.symp.ser.225-232)。可選擇地,可以利用合成多肽氨基酸序列或其一部分的方法來產生蛋白本身。例如,可以利用多種固相技術進行肽合成(roberge,j.y.etal.(1995)science269:202-204),并且例如利用abi431a肽合成儀(perkinelmer,paloalto,ca)可以實現自動化合成。
可以利用制備型高效液相色譜(例如,creighton,t.(1983)proteins,structuresandmolecularprinciples,whfreemanandco.,newyork,n.y.)或領域內可用的其它類似技術來基本上純化新合成的肽。可以通過氨基酸分析或測序(例如,edman降解法)來確認合成肽的組成。此外,在直接合成和/或利用化學方法與來自其它蛋白的序列或序列的任何部分進行組合的過程中,多肽的氨基酸序列或其任何部分可被改變,從而產生變體多肽。
為了表達所需的多肽,可以將編碼多肽的核苷酸序列或功能等同物插入合適的表達載體中,即包含插入的編碼序列的轉錄和翻譯所必需的元件的載體。可以用本領域技術人員公知的方法構建含有編碼目的多肽的序列和合適的轉錄和翻譯控制元件的表達載體。這些方法包括:體外重組dna技術、合成技術和體內遺傳重組。這些技術例如描述于sambrook,j.etal.(1989)molecularcloning,alaboratorymanual(分子克隆,實驗手冊)和ausubeletal.(2001-2008currentprotocolsinmolecularbiology(現代分子生物學技術),greenepubl.assoc.inc.&johnwiley&sons,inc.,ny,ny)。
多種表達載體/宿主系統可以用來包含和表達多核苷酸序列。這些表達載體/宿主系統包括但不限于,諸如用重組噬菌體、質粒、或粘粒dna表達載體轉化的細菌的微生物;用酵母表達載體轉化的酵母;用病毒表達載體(例如,桿狀病毒)感染的昆蟲細胞系統;用病毒表達載體(例如,花椰菜花葉病毒,camv;煙草花葉病毒tmv)或用細菌表達載體(例如,ti或pbr322質粒)轉化的植物細胞系統;或動物細胞系統。
表達載體中存在的“控制元件”或“調節序列”是與宿主細胞蛋白相互作用來進行轉錄和翻譯的載體的那些非翻譯區(增強子、啟動子、5'和3'非翻譯區)。這些元件在它們的強度和特異性上可以不同。根據所用的載體系統和宿主,可以使用任何數量的合適的轉錄和翻譯元件,包括組成型啟動子和誘導型啟動子。例如,當克隆入細菌系統時,可以使用誘導型啟動子,諸如pbluescript噬菌粒(stratagene,lajolla,calif.)或psport1質粒(gibcobrl,gaithersburg,md.)的雜合lacz啟動子等。在哺乳動物細胞系統中,通常優選來自哺乳動物基因或來自哺乳動物病毒的啟動子。如果有必要產生含有編碼多肽的序列的多拷貝的細胞系,則可以優選地使用基于sv40或ebv的、帶有合適的選擇標記的載體。
在細菌系統中,根據表達的多肽的希望的用途,可以選擇多種表達載體中的任何一種。例如,當大量需要時,例如用于誘導抗體時,可以使用指導容易純化的融合蛋白的高水平表達的載體。這類載體包括但不限于,諸如pbluescript(stratagene)的多功能大腸桿菌克隆和表達載體,其中將編碼目的多肽的序列與載體連接,與氨基末端met和隨后的β-半乳糖苷酶的7個殘基在同一讀碼框中,以便產生雜合蛋白;pin載體(vanheeke,g.ands.m.schuster(1989)j.biol.chem.264:5503-5509)等。pgex載體(promega,madison,wis.)也可以用來將外源多肽表達為具有谷胱甘肽巰基轉移酶(gst)的融合蛋白。通常,這類融合蛋白是可溶的,并且可以通過谷胱甘肽瓊脂糖珠的吸附并隨后在游離谷胱甘肽的存在下洗脫來容易地從裂解的細胞中純化。這些系統中產生的蛋白可以被設計為包括肝素、凝血酶或因子xa蛋白酶切割位點,以便任意地從gst部分釋放克隆的目的多肽。
在釀酒酵母(saccharomycescerevisiae)中,可以使用多種含有諸如α因子、醇氧化酶和pgh的組成型或誘導型啟動子的載體。對于綜述而言,可以參見ausubeletal.(同上)和grantetal.(1987)methodsenzymol153:516-544。
如果使用植物表達載體,編碼多肽的序列的表達可以由多種啟動子中的任何一種所驅動。例如,諸如camv的35s和19s啟動子的病毒啟動子可以單獨使用或與來自tmv的ω前導序列聯合使用(takamatsu,n.(1987)emboj.6:307-311)。可選擇地,可以使用諸如rubisco的小亞基或熱休克啟動子的植物啟動子(coruzzi,g.etal.(1984)emboj.3:1671-1680;broglie,r.etal.(1984)science224:838-843;以及winter,j.etal.(1991)resultsprobl.celldiffer.17:85-105)。可以通過直接的dna轉化或病原體介導的轉染將這些構建體導入植物細胞。這類技術描述于很多可普遍獲得的綜述中(參見例如,hobbs,s.ormurry,l.e.inmcgrawhillyearbookofscienceandtechnology(1992)mcgrawhill,newyork,n.y.;pp.191-196)。
還可以用昆蟲系統來表達目的多肽。例如,在一種這類系統中,將苜蓿銀紋夜蛾核型多角體病毒(acnpv)用作載體在草地貪夜蛾(spodopterafrugiperda)細胞或粉紋夜蛾(trichoplusia)幼蟲中表達外來基因。可以將編碼多肽的序列克隆入病毒的非必需區,例如多角體蛋白基因,并置于多角體蛋白啟動子的控制下。編碼多肽的序列的成功插入會使多角體蛋白基因失活,并產生缺少衣殼蛋白的重組病毒。隨后,可以將所述重組病毒用于感染例如草地貪夜蛾細胞或粉紋夜蛾幼蟲,其中目的多肽可以表達(engelhard,e.k.etal.(1994)proc.natl.acad.sci.91:3224-3227)。
在哺乳動物宿主細胞中,通常可以使用多種基于病毒的表達系統。例如,如果將腺病毒用作表達載體,則可以將編碼目的多肽的序列連入由晚期啟動子和三聯前導序列組成的腺病毒轉錄/翻譯復合物中。病毒基因組的非必需e1或e3區中的插入可以用來獲得能在感染的宿主細胞中表達多肽的活病毒(logan,j.andshenk,t.(1984)proc.natl.acad.sci.81:3655-3659)。此外,可以使用諸如魯斯氏肉瘤病毒(rsv)增強子的轉錄增強子來增強哺乳動物宿主細胞中的表達。
也可以使用特異的起始信號來實現編碼目的多肽的序列的更高效的翻譯。這些信號包括atg起始密碼子和鄰近的序列。如果將編碼多肽的序列、其起始密碼子和上游序列插入合適的表達載體中,則可以不需要其他的轉錄或翻譯控制信號。然而,如果僅將編碼序列或其一部分插入,則應該提供包括atg起始密碼子在內的外源翻譯控制信號。此外,起始密碼子應該在正確的讀碼框中以確保全部插入物的翻譯。外源翻譯元件和起始密碼子可以是不同來源的,可以是天然的或合成的。可以通過包含適于所用的特定細胞系統的增強子來增強表達效率,諸如文獻中描述的那些增強子(scharf,d.etal.(1994)resultsprobl.celldiffer.20:125-162)。
此外,可以基于宿主細胞株調節插入的序列的表達或以所需的方式加工所表達的蛋白的能力來選擇宿主細胞株。多肽的這些修飾包括但不限于,乙酰化、羧基化、糖基化、磷酸化、脂質化和酰基化。切割蛋白的“前原”形式的翻譯后加工也可以用來促進正確的插入、折疊和/或功能。可以選擇諸如cho、cos、hela、mdck、hek293和wi38的、對于這類翻譯后活性具有特異性的細胞體系和特征機制的不同宿主細胞,以確保外源蛋白的正確修飾和加工。
為了長期、高產量地產生重組蛋白,通常優選穩定的表達。例如,可以用表達載體轉化穩定表達目的多核苷酸的細胞系,所述表達載體可以在同一載體或分別的載體上含有病毒復制起點和/或內源表達元件以及選擇標記基因。引入載體后,在將細胞轉至選擇培養基之前,可以允許細胞在富集培養基中生長1-2天。選擇標記的目的是賦予對于選擇的抗性,并且選擇標記的存在允許成功表達引入的序列的細胞的生長和恢復。可以用適合細胞類型的組織培養技術增殖穩定轉化的細胞的抗性克隆。
可以用任何數量的選擇系統來恢復轉化的細胞系。這些選擇系統包括但不限于,可以分別在tk-細胞或aprt-細胞中使用的單純皰疹病毒胸苷激酶(wigler,m.etal.(1977)cell11:223-32)和腺嘌呤磷酸核糖基轉移酶(lowy,i.etal.(1990)cell22:817-23)。此外,抗代謝物、抗生素或除草劑抗性可以被用作選擇的基礎;例如,賦予對氨甲喋呤的抗性的dhfr(wigler,m.etal.(1980)proc.natl.acad.sci.77:3567-70);賦予對氨基糖苷類、新霉素和g-418的抗性的npt(colbere-garapin,f.etal(1981)j.mol.biol.150:1-14);以及分別賦予對氯磺隆和草胺磷乙酰轉移酶的抗性的als或pat(murry,同上)。其它選擇基因已有描述,例如,允許細胞利用吲哚來代替色氨酸的trpb,或允許細胞利用組胺醇來代替組氨酸的hisd(hartman,s.c.andr.c.mulligan(1988)proc.natl.acad.sci.85:8047-51)。可視標志物已獲得普遍應用,這類標志物如花青素、β-葡糖苷酸酶及其底物gus、以及熒光素酶及其底物熒光素,這類標記不僅廣泛用于鑒定轉化株,而且用于對歸因于特異性載體系統的瞬時或穩定蛋白表達進行定量(rhodes,c.a.etal.(1995)methodsmol.biol.55:121-131)。
盡管標志基因表達的存在與否能提示同時存在目的基因,但是標志基因的存在和表達需要被證實。例如,如果將編碼多肽的序列插入標志基因序列中,則可以通過標志基因功能的缺失來鑒定含有序列的重組細胞。可選擇地,可以將標志基因與編碼多肽的序列串聯置于一個啟動子的控制下。標志基因響應于誘導或選擇的表達通常能同時表明串聯基因的表達。
可選擇地,可以通過本領域技術人員已知的多種方法來識別含有并表達所需多核苷酸序列的宿主細胞。這些方法包括但不限于,dna-dna或dna-rna雜交和蛋白生物測定或免疫測定技術,蛋白生物測定或免疫測定技術包括例如,用于對核酸或蛋白進行檢測和/或定量的基于膜、溶液或芯片的技術。
使用對產物特異的多克隆或單克隆抗體檢測和測量多核苷酸編碼的產物的表達的多種方法在領域內是已知的。實例包括酶聯免疫吸附測定(elisa)、放射性免疫測定(ria)和熒光活化細胞分選(facs)。利用與給定多肽上的兩個非干擾表位起反應的單克隆抗體的基于雙點、單克隆的免疫測定對于某些應用可以是優選的,但是還可以利用競爭性結合測定。這些測定和其他測定描述于hampton,r.etal.(1990;serologicalmethods,alaboratorymanual(血清學方法:實驗手冊),apspress,stpaul.minn.)和maddox,d.e.etal.(1983;j.exp.med.158:1211-1216)等中。
廣泛多種的標記物和綴合技術是本領域技術人員已知的并且可以用于各種核酸和氨基酸測定中。產生用于檢測多核苷酸相關序列的標記的雜交或pcr探針的手段包括寡核苷酸標記、缺口平移、末端標記或使用標記的核苷酸的pcr擴增。可選擇地,可以將序列或其任何部分克隆入載體中,用于產生mrna探針。這類載體在本領域內是已知的并且是可商業購得的,并且可以通過添加諸如t7、t3或sp6的合適的rna聚合酶和標記的核苷酸來體外合成rna探針。可以使用多種可商業購得的試劑盒進行這些步驟。可以使用的合適的報告分子或標記包括放射性核素、酶、熒光劑、化學發光劑或產色劑以及底物、輔因子、抑制劑、磁性顆粒等。
可以在適合蛋白從細胞培養物中表達和回收的條件下,培養用目的多核苷酸序列轉化的宿主細胞。根據序列和/或所用的載體,重組細胞產生的蛋白可以是分泌的或包含在細胞內。如本領域技術人員所理解地,可以將包含本發明多核苷酸的表達載體設計為包含指導編碼的多肽通過原核生物或真核生物細胞膜的分泌的信號序列。可以用其它的重組構造,將編碼目的多肽的序列與編碼有助于可溶蛋白純化的多肽結構域的核苷酸序列連接起來。這類有助于純化的結構域包括但不限于,允許對固定化金屬進行純化的諸如組氨酸-色氨酸模塊的金屬螯合肽、允許對固定化免疫球蛋白進行純化的蛋白a結構域和在flags延伸/親和純化系統中所用的結構域(immunexcorp.,seattle,wash.)。在純化結構域和所編碼的多肽之間包含可切割連接物序列,諸如對因子xa或腸激酶特異的那些序列(invitrogen.sandiego,calif.),可用于促進純化。一種這類表達載體提供了這樣的融合蛋白的表達,該融合蛋白含有目的多肽和編碼6個組氨酸殘基的核酸,以及位于核酸之后的硫氧還蛋白或腸激酶切割位點。如在porath,j.etal.(1992,prot.exp.purif.3:263-281)中所述,組氨酸殘基有助于在imiac(固定化金屬離子親和層析)上進行純化,而腸激酶切割位點提供了一種從融合蛋白純化所需多肽的方法。對含有融合蛋白的載體的討論提供在kroll,d.j.etal.(1993;dnacellbiol.12:441-453)中。
除了重組產生方法,可以通過使用固相技術的直接肽合成來產生本發明的多肽及其片段(merrifieldj.(1963)j.am.chem.soc.85:2149-2154)。可以用手動技術或通過自動化來進行蛋白合成。例如,可以用appliedbiosystems的431a肽合成儀(perkinelmer)實現自動化合成。可選擇地,可以分別地化學合成不同片段并使用化學方法將片段組合來產生全長分子。
耐受
免疫耐受被定義為對抗原,通常是參與引起疾病的抗原的免疫無應答。盡管耐受可以通過不同途徑給予抗原來誘導,但是口服耐受是指抗原的口服給藥,這已經在數種動物模型中導致疾病活動的抑制,包括實驗自身免疫性腦脊髓炎——多發性硬化的嚙齒動物模型、重癥肌無力、葡萄膜炎、胰島素依賴型糖尿病和膠原誘導的關節炎(fariaandweiner1999)。來自人類臨床試驗的早期結果表明,口服耐受在自身免疫性葡萄膜炎、糖尿病、鎳過敏癥以及可能在多發性硬化中都是有效的(fariaandweiner1999;dudaetal.2000)。有數項研究報道了在器官移植中誘導口服耐受(sayeghetal.1996;hancocketal.1993;ishidoetal.1999;sayeghetal.1992a;sayeghetal.1992b)。在每篇報道中,通過在移植之前食用供體mhc來源的肽或食用異源細胞來誘導耐受(sayeghetal.1996;hancocketal.1993;ishidoetal.1999;sayeghetal.1992a;sayeghetal.1992b)。這些技術可有效預防對心臟和角膜同種移植物的排斥(sayeghetal.1996;hancocketal.1993;ishidoetal.1999;sayeghetal.1992a;sayeghetal.1992b;fariaandweiner1999)。在這些研究中,除了降低疾病活動之外,由口服耐受所誘導的免疫抑制還可以通過響應于靶抗原的遲發型超敏反應(dth)的下調以及細胞免疫和體液免疫的減弱來衡量(fariaandweiner1999;mayer2000;garsideandmowat1997)。
口服(和其它給藥途徑)耐受通過以下三種機制下調抗原特異的免疫應答:1.抗原特異性細胞的活性抑制、2.抗原特異性細胞的克隆無反應和3.抗原特異性細胞的克隆排除(fariaandweiner1999,milleretal.1991;chenetal.1994;chenetal.1995)。盡管所有三種機制可以在響應于口服耐受時同時起作用,但是活性抑制和克隆無反應是口服耐受所誘導的免疫抑制的關鍵機制(fariaandweiner1999)。
活性抑制描述的是一種淋巴細胞亞類以抗原特異的方式受到另一種物質的調節。根據抗原和疾病狀態,抑制細胞可以是cd4+和/或cd8+t淋巴細胞,其從諸如脾或周圍淋巴結的外周淋巴組織遷移至疾病活動位點。將這些細胞過繼轉移至天然接受者已經證實這些細胞對卵白蛋白誘導的超敏性和多發性硬化的嚙齒動物模型中活性抑制的作用。由以下數據證實活性抑制的體外證據,來自動物的耐受淋巴細胞能跨越transwell細胞培養系統抑制其它抗原特異性t淋巴細胞的增殖(fariaandweiner1999;milleretal.1991)。
克隆無反應是指抗原特異性t淋巴細胞的無應答,其特征為暴露于抗原之后增殖減弱,并且參與數種動物模型的口服耐受。無反應可能是由cd4+或cd8+t淋巴細胞自身、局部環境中其它t淋巴細胞或細胞所產生的可溶性抑制因子的結果,或者是因為合適輔刺激分子的表達降低(fariaandweiner1999)。克隆排除是指去除抗原特異性t淋巴細胞,但是很少有報道是作為抗原口服耐受的機制(chenetal.1995)。
在口服耐受期間抑制免疫應答的可溶性調節物主要來源于調節性或抑制性t淋巴細胞(fariaandweiner1999)。有五類t淋巴細胞通過其產生的細胞因子進行描述:產生白細胞介素-2(il-2)和γ干擾素(γifn)的th1型;產生il-4和il-10的th2型;單獨產生高水平的轉化生長因子β(tgf-β)或聯合產生非常低水平的il-4、il-10或γifn的th3型;產生高水平的il-10和低水平的tgf-β的tr1細胞(fariaandweineretal.1999;mayer2000;garsideandmowat1997;grouxetal.1997);以及產生il-17的th17細胞(參見例如,immunolrev.2008,226:87-102;nature.2006may11;441(7090):235-8)。由于th3、th2和tr1-t淋巴細胞已被證實是口服耐受所誘導的活性抑制的主要調節物,所以tgf-β、il-4和il-10被認為是該過程中的關鍵細胞因子(tengetal.1998;shietal.1999b)。來自baroneetal.等顯示在不存在這些細胞因子下所發生的口服耐受誘導的報道提示,其它調節物或細胞也能抑制免疫應答(baroneetal.1998;shietal.1999a)。
盡管針對口服耐受的研究集中在抑制免疫應答的t淋巴細胞來源的細胞因子,但是并非由t淋巴細胞產生的一氧化氮已知是同種免疫應答的有效抑制劑(garsideandmowat1997)。表明一氧化氮調節凋亡(其參與排斥應答)的這些數據和其它數據(meyeretal.1998;kallioetal.1997;shiraishietal.1997;shiraishietal.1995;medot-pirenneetal.1999)提示一氧化氮可能是口服耐受的調節物并且阻止排斥應答。tgf-β是一氧化氮合成的有效誘導物,并且是口服耐受中活性抑制的關鍵調節物(fariaandweiner1999;meyeretal.1998;vodovotzetal.1998;vodovitzetal.1999)。因此,在耐受宿主中由tgf-β誘導的免疫抑制在一定程度上可以由一氧化氮調節。然而,一氧化氮響應于口服耐受的產生是未知的。
由apc誘導的抗原特異性t淋巴細胞的激活需要t淋巴細胞和apc之間的雙向相互作用。最初,apc呈遞與t細胞受體結合的mhc分子,這刺激t淋巴細胞上cd40-配體(cd40l)表達的上調。cd40l轉而與其受體,位于apc上的cd40結合。通過cd40的信號轉導誘導apc上cd80和cd86的表達,它們一旦與其受體,t淋巴細胞上的cd28結合,就會導致共刺激和隨后的t淋巴細胞激活(liuetal.1999;lietal.1999;ledermanandsiciu-foca1999)。盡管對口服耐受誘導的研究集中在t淋巴細胞功能上,最近來自taamsetal(1998)的研究報道了耐受誘導可以影響apc的功能,這與其他研究者的數據相似(wuetal.1998;finkelmanetal.1996;vineyetal.1998)。例如,wu.etal(1998)的報道顯示來自口服耐受小鼠淋巴結和脾的apc上cd80的表達降低,這提示無效的apc可有助于耐受接受者中受損t淋巴細胞的激活。此外,體外研究顯示抑制性t淋巴細胞能抑制apc中cd86的表達,這突出耐受如何誘導受損apc功能的另一機制(liuetal.1999;lietal.1999;ledermanandsiciu-foca1999)。
col(v)的給予除了能防止對自身的增殖應答之外,還能防止對同種抗原的增殖應答,并且能防止接受者肺部急性排斥病理的發展(參見例如,美國專利第7,348,005號;wo2007/120947)。因此,col(v)可以誘導對供體同種抗原和對自身的無反應,或可選擇地,對供體抗原和對colv的增殖應答的缺乏可能是由于同種抗原特異性肺淋巴細胞的克隆排除;或者可選地,可能是由于抑制細胞的活性。
不希望受理論的束縛,由colv誘導的耐受可以通過相關的抑制進行誘導(參見例如,humimmunol.2008nov;69(11):715-20)。而且,認為colv與淋巴細胞上膠原受體的差異結合導致可以涉及差異激活信號(參見例如,cellsignal.2006aug;18(8):1108-16)。例如,只有v型膠原誘導t細胞中的il-17信號轉導。
抗原的口服給藥是誘導周圍t細胞耐受性的有效方法。這種現象通常被稱為口服耐受,已經在多種動物自身免疫疾病模型中進行了良好的研究,包括腦脊髓炎、葡萄膜炎、糖尿病、重癥肌無力和關節炎。然而,對誘導耐受的機制還不完全了解。所有已知的耐受誘導機制,包括克隆無反應、克隆排除和受il-4、il-10或tgf-β調節介導的活性抑制都可以在口服耐受中發揮作用(fariaandweiner,1999)。通常,較高劑量的抗原被報道能誘導無反應或克隆排除(chenetal.,1995;whitacreetal.,1991),而低劑量能誘導細胞因子調節和活性抑制(fariaandweiner,1999;chenetal.,1994)。在心臟移植的動物模型中,同種脾細胞的口服給藥被證實在旁路th1激活和選擇性刺激誘導th-2來源的抑制性細胞因子(例如il-4)的耐受誘導中是有效的(hancocketal.,1993;ishidoetal.,1999)。
因此,口服耐受是通過向個體口服給藥抗原(即通過食用)來下調免疫應答的方法。口服耐受的特征為系統性抗體產生的水平降低以及遲發型超敏反應(dth)、t細胞增殖、細胞毒性反應和移植排斥減弱(alpanetal.2001.j.immunol.166:4843-52;chenetal.1995.nature376:177-80;weiner.1997.imm.today.7:335-44;sayeghetal.1992.transplantation.53:163-6)。
本文還考慮用于誘導耐受的其它途徑,尤其是通過肌肉內注射、皮下注射、皮內注射和靜脈內注射。本文在低劑量易于誘導耐受和高劑量能誘導免疫應答的情況下考慮皮內注射。s
對耐受的研究主要集中在耐受性抗原對t淋巴細胞功能的影響和t淋巴細胞在抑制免疫激活中的作用上(fariaandweiner1999;mayer2000;garsideandmowat1997)。然而,對任何抗原的免疫應答需要apc和t淋巴細胞之間的相互作用,并且t淋巴細胞可以影響apc的功能(liuetal.1999;lietal.1999;ledermanandsiciu-foca1999)。因此,由來自耐受宿主的apc所呈遞的下調的抗原對于耐受誘導的貢獻可以間接地由于與抑制性t淋巴細胞的相互作用,或者可能由于耐受抗原對apc的直接影響。
因此,在本發明的某些方面中,本發明提供了恢復或增強copd和哮喘患者對colv的自我耐受的方法。一個實施方案是通過口服療法給予colv(yasufukuetal,2001;yasufuku,etal,2002)使其通過間隙進入肺中或通過給藥方案上的其它脫敏策略來治療copd或哮喘的方法,所述給藥方案被設計成用于增強患者對膠原(包括但不必要限制于colv及其抗原性成分和變體)的耐受性。
“耐受原性片段”表示能誘導對全長蛋白的耐受性的片段,該片段是所述全長蛋白的片段(例如,全長v型膠原及其耐受原性片段)。在某些實施方案中,耐受原性片段至少可以誘導對全長v型膠原的耐受性,以及全長v型膠原蛋白可以并且在某些實施方案中在誘導耐受性上可以比全長膠原蛋白更加有效。然而,在某些實施方案中,耐受原性片段誘導對全長v型膠原的耐受性,但是可以不會與全長v型膠原蛋白同樣有效地誘導耐受性。這種耐受原性片段仍然可以用在本發明中,尤其當所述耐受原性片段具有其它優勢時,例如與全長蛋白相比便制備或純化。如同本領域技術人員所認識到的,多種已知的測定可用于評價耐受性的誘導,包括檢測延遲型超敏反應(dth)應答、通過elisa或其它方法檢測細胞因子產生、t細胞增殖或細胞毒性測定、b細胞增殖測定、抗體產生等。這類測定是領域內已知的,并且例如描述于currentprotocolsinimmunology(現代免疫學技術),editedby:johne.coligan,adam.kruisbeek,davidh.margulies,ethanm.shevach,warrenstrober(2001johnwiley&sons,ny,ny);ausubeletal.(2001currentprotocolsinmolecularbiology(現代分子生物學技術),greenepubl.assoc.inc.&johnwiley&sons,inc.,ny,ny);美國專利第7,348,005號中以及其它地方。
因此,耐受原性片段是諸如v型膠原或其任何一種或多種α鏈的耐受原性多肽的片段,該片段本身對于通過b細胞和/或t細胞的表面受體(例如,b細胞抗體受體或t細胞受體)來識別多肽的特異性b細胞和/或t細胞而言在免疫學上是耐受原性的(即誘導耐受性)。通常利用公知的技術來鑒定耐受原性片段,例如paul,fundamentalimmunology,3rded.,(ravenpress,1993)中以及其中所引用的參考文獻中所總結的那些技術。這類技術包括篩查有能力降低t細胞和/或b細胞反應性的多肽。如本文所用,如果抗血清和抗體與抗原特異結合(即它們與elisa或其它免疫測定中的蛋白特異性地反應并且與不相關的蛋白不發生可檢測到的反應),則它們是“抗原特異性的”。可以按本文所述和利用公知技術來制備這類抗血清和抗體。
在一個實施方案中,本發明多肽的耐受原性片段是以基本上不低于全長多肽的耐受原活性的水平誘導b細胞和/或t細胞耐受性的一部分(例如,在諸如抗體產生的合適的測定中,可以通過elisa檢測和/或t細胞反應性測定(t細胞增殖或細胞因子產生測定)。優選地,耐受原性部分的耐受原活性水平為全長多肽耐受原活性的至少約50%、55%、60%、65%、70%、75%、80%、85%或高于約90%、91%、92%、93%、94%、95%、96%、97%、98%或99%。在一些實例中,鑒定耐受原活性水平大于對應全長多肽耐受原活性水平的耐受原性片段,例如,具有高于約100%、110%、120%、130%、140%或150%或更高的耐受原活性。
在某些實施方案中,可以利用計算機分析鑒定耐受原性片段,例如tsites程序(參見rothbardandtaylor,emboj.7:93-100,1988;deavinetal.,mol.immunol.33:145-155,1996),其搜索具有激發th應答潛能的肽基序。可以按照bimas(parkeretal.,j.immunol.152:163,1994)和其它hla肽結合預測分析來鑒定具有適于結合鼠和人i類或ii類mhc的基序的ctl肽。可選擇地,可以利用確定的肽結合基序來鑒定與特定mhc分子結合的部分,例如rammenseeetal.,immunogenetics41:178-228,1995中所描述的那些基序。為了確認與鼠和人i類或ii類mhc分子結合的肽,可以使用領域內已知的肽結合測定。為了證實免疫原性或耐受原性,可以利用hlaa2或其它轉基因小鼠模型和/或利用樹突細胞、成纖維細胞或外周血細胞的體外刺激測定來測試肽。
應當指出,在某些實施方案中,本發明的耐受原性片段還是免疫原性片段。在這方面中,如同本領域技術人員應當認識到的,高免疫原性片段,例如類似于colv的蛋白的優勢免疫表位,當正確給予時(例如,通常以低劑量持續一段時間)可以是耐受原性的。因此,本發明還考慮colv的免疫原性片段的鑒定和使用,其中這類免疫原性片段可用來誘導耐受性。在這方面中,免疫原性部分的免疫原活性水平為全長多肽免疫原活性的至少約50%、55%、60%、65%、70%、75%、80%、85%或大于約90%、91%、92%、93%、94%、95%、96%、97%、98%或99%。在一些實例中,鑒定免疫原活性水平大于對應全長多肽免疫原活性水平的免疫原性片段,例如,具有高于約100%、110%、120%、130%、140%或150%或更高的免疫原活性。
用來鑒定免疫原性片段的分析可以與用來鑒定耐受原性片段的分析一樣,包括利用諸如tsites程序(參見rothbardandtaylor,emboj.7:93-100,1988;deavinetal.,mol.immunol.33:145-155,1996)的計算機分析,所述tsites程序搜索具有激發th應答潛能的肽基序。可以按照bimas(parkeretal.,j.immunol.152:163,1994)和其它hla肽結合預測分析來鑒定具有適于結合鼠和人i類或ii類mhc的基序的ctl肽。可選擇地,可以利用確定的肽結合基序來鑒定與特定mhc分子結合的部分,例如rammenseeetal.,immunogenetics41:178-228,1995中所描述的那些基序。為了確認與鼠和人i類或ii類mhc分子結合的肽,可以使用領域內已知的肽結合測定。為了證實免疫原性或耐受原性,可以利用hlaa2或其它轉基因小鼠模型和/或利用樹突細胞、成纖維細胞或外周血細胞的體外刺激測定來測試肽。
本發明的方法中考慮使用完整的v型膠原或其具有耐受原或免疫原活性的成分α鏈的任何一種或多種。因此,v型膠原的耐受原性片段或免疫原性片段可以指完整v型膠原的片段或可以指任何一種成分α鏈的耐受原性或免疫原性片段。在某些實施方案中,本文所用的colv可以包含由三種α鏈組成的膠原分子。在其它實施方案中,colv可以包含α鏈的任何一種或多種,例如由seqidno:1、3或5所示的多核苷酸所編碼的seqidno:2、4或6所示的那些α鏈或這些α鏈的耐受原性片段或免疫原性片段。
在某些實施方案中,兩種或更多種耐受原性或免疫原性片段可以被同時使用、單獨給予、混在組合物中或作為融合蛋白。在這方面中,任何數量的耐受原性片段或免疫原性片段可用來誘導對colv的耐受性,例如2、3、4、5、6、7、8、9、10或更多種耐受原性或免疫原性片段作為單獨的片段處于組合物中或作為融合蛋白(具有或不具有連接物)。在某些實施方案中,本發明的方法可以使用11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30或更多種片段。
按照某些實施方案,可以利用多種免疫測定方法中的任何一種,例如通過elisa方法來完成對抗膠原抗體存在的檢測。如同通過參考標準免疫測定教科書所看到的,可以利用大量的免疫測定技術,這些包括但不限于非競爭類型的單點和雙點或“夾心”測定以及傳統的競爭結合測定。
夾心測定是最有用且常用的基于抗體的測定方法,并且可用來實施多個實施方案。夾心測定技術存在大量變型,并且所有都意圖包括在各實施方案中。簡單而言,在用于檢測樣品中的抗體的典型測定中,將非標記的抗原固定在固相基底上,并將待檢驗的樣品與結合的抗原分子接觸。在一段合適的孵育時間,即持續足以允許形成抗體-抗原復合物的時間段之后,添加諸如抗人igg的二抗(標記有能產生檢測信號的報告分子),并孵育,允許足以形成抗體-抗原-標記抗體的時間。洗掉任何未反應的物質,并通過觀察所述報告分子所產生的信號來測定樣品中待檢測的抗體的存在。例如通過簡單觀察可視信號結果可以是定性的,或者通過將目的樣品所產生的信號與含有已知量的待檢測抗體的對照樣品進行比較,可以對結果進行定量。這種測定的變型包括同時測定,其中樣品和標記抗體被同時添加到結合的抗原上。這些技術是公知的,包括本領域技術人員顯而易見的任何微小變型。在典型的夾心測定中,抗原是固定的,例如被共價或被動地連接于固相表面。在一些實施方案中,所述固相表面通常為玻璃或聚合物,最常用的聚合物是纖維素、聚丙烯酰胺、尼龍、聚苯乙烯、聚氯乙烯或聚丙烯。固相支持體可以是管、珠、盤或微型板的形式或適于進行免疫測定的其它任何表面。多種結合方法是領域內已知的,并且通常由抗原與給定表面的交聯、共價結合或物理吸附組成。然后在用于添加測驗樣品的配制液中洗滌固定的抗原。然后將待測驗的樣品等分與固定的抗原接觸,并在合適的條件(例如25℃)下孵育足以允許任何抗體與樣品中存在的膠原結合的時間段(例如2-40分鐘)。接觸時間的實際長度、緩沖液條件、溫度等是易于調整的參數,并且對于給定的測驗通常是容易獲得的。孵育之后,洗滌包含任何結合抗體的固定的抗原并干燥,并與所述結合抗體特異的二抗一起孵育,所述二抗例如抗人igg。二抗與用來指示二抗與抗體-固定的抗原復合物的結合的報告分子相連。
一種用于檢測抗colv抗體的特定方法描述于wo2007/120947中。具體而言,該珠子測定檢測抗v型膠原抗體,所述抗體可以存在于患自身免疫應答v型膠原的患者的血清和/或肺灌洗液中。為該項測定提供v型膠原包被的珠子和其它必需試劑。簡單而言,典型測定如下:1)用無菌pbs洗滌鏈霉親和素包被的珠子(例如,來自于polyscience,warrington,pa的那些)。并將珠子懸浮于合適體積的具有人v型膠原的pbs中。2)用抗人v型膠原抗體的兔抗體(bioten)(abeam,cambridge,ma),按照上述1中相同的方法產生陽性對照。3)對于每次測定,將偶聯的珠子在pbs中洗滌,并在pbs加肺灌洗液漿液中進行孵育。然后用含有10%fcs的pbs洗滌珠子。4)然后將珠子懸浮于無菌pbs+10%fbs中,并在室溫下與二抗一起孵育。通常,使用與r-pe偶聯的抗人igg抗體(sigma,saintlouis),但是如同本領域技術人員應當理解的,可以使用其它合適的抗體。在這方面中,在某些實施方案中,為了檢測疾病進展中從一種亞型向另一種的轉變,可以使用抗人igg1-、igg2-、igg3-或igg4-特異性抗體。將珠子在含有10%fcs的pbs中洗滌,懸浮于pbs/fcs溶液中,并利用流式細胞儀進行分析。對于陽性對照,可以向珠子測定中添加已知量的抗colv的抗血清或抗體。
通常,見于copd和哮喘患者中的抗colv抗體是igg,但是也可以存在其它抗體類型,例如igm。此外,在本發明的某些實施方案中,抗v型膠原的igg抗體亞型隨疾病進展發生改變,所述v型膠原可以存在于患者的血清和/或肺灌洗液中。在這方面中,igg亞型在疾病進展中可以發生轉變,并且某些亞型可以指示疾病在惡化。因此,在疾病進展中可以存在任何一種或多種igg亞型,例如igg1、igg2、igg3或igg4或以上的任何組合。本發明提供了利用本文所述的珠子測定檢測v型膠原特異的igg1、igg2、igg3、igg4亞型抗體的方法。如同本領域技術人員應當認識到的,igg亞型特異的抗體是可購買得到的,并且可用于本文所述的方法中。在某些實施方案中,igg1的增加指示疾病在惡化。在另一實施方案中,轉變成igg2亞型指示疾病在惡化。在另一實施方案中,轉變成igg3亞型指示疾病在惡化。在另一實施方案中,轉變成igg4亞型指示疾病在惡化。
組合物、藥物組合物和使用方法
可以通過用于提供相似功效的藥劑的任何可接受的給藥方式,以純形式或合適的藥物組合物來實施本發明的耐受原性化合物或組合物或其藥學可接受的鹽的給藥。如本文其它地方所指出的,本文所考慮的一種用于誘導耐受性的途徑是口服給藥。然而,還可以使用多種其它途徑中的任何一種,尤其包括靜脈內注射、肌肉內注射、皮內注射、皮下注射以及其它途徑。可以通過將本發明的化合物與合適的藥學可接受的載體、稀釋液或賦形劑組合來制備本發明的藥物組合物,并且可以配制成固體、半固體、液體或氣態形式的制劑,例如片劑、膠囊、粉末、顆粒、軟膏、溶液、栓劑、注射劑、吸入劑、凝膠、微球和氣霧劑。此外,其它藥學活性組分(包括其它免疫抑制劑)和/或諸如鹽、緩沖液和穩定劑的合適的賦形劑可以,但不需要存在于組合物中。
可以通過多種不同的途徑來實現給藥,包括口服、腸胃外、鼻、靜脈內、皮內、皮下或局部。給藥的優選方式取決于待治療或預防的疾病狀態的性質。給藥后,能降低、抑制、防止或延遲抗colv免疫應答或這類應答的臨床適應癥的發生的量被認為是有效的。
在某些實施方案中,所給予的量足以導致本文其它地方所述的免疫活性的降低(例如,t細胞應答、b細胞應答、抗colv抗體的水平等)。治療的精確劑量和持續時間是待治療疾病的函數,并且可以使用已知的測驗方案通過經驗來確定或者通過在領域內已知的模型系統中測驗組合物并外推來確定。還可以進行可控的臨床試驗。劑量還可以隨待緩解的疾病狀態的嚴重程度而改變。藥物組合物通常被配制和給藥以發揮治療有用的效應同時降低不需要的副作用的。組合物可以一次給藥,或可以分成多個更小的劑量分多次給藥。對于任何具體的患者,可以按照患者需求隨時間調整具體的劑量方案。
本發明的組合物可以單獨給藥或與其它已知的治療結合,例如免疫抑制方案、放射療法、化學療法、移植、口服膠原療法、免疫療法、激素療法、光動力療法。
給予這些組合物和相關藥物組合物的典型途徑因而包括但不限于,口服、局部、經皮、吸入、肺內灌注、腸胃外、舌下、口腔、直腸、陰道和鼻內途徑。本文所用的術語腸胃外包括皮下注射、靜脈內注射、肌肉內注射、胸骨內注射或灌輸技術。本發明的組合物被配制成一旦將該組合物給予患者時允許其中所含的活性成分可為生物所利用。被給予個體或患者的組合物采用一或多劑量的形式,其中例如,片劑可以是單劑量單位,且氣霧劑形式的本發明化合物的容器可以持有多個劑量單位。制備這類劑量形式的實際方法是已知的,或者對本領域技術人員而言是顯而易見的;例如,參見remington:thescienceandpracticeofpharmacy,20thedition(philadelphiacollegeofpharmacyandscience,2000)。在任何情況下,按本發明的教導,待給予的組合物含有治療有效量的本發明的化合物或其藥學上可接受的鹽,用于治療感興趣的疾病或疾病狀態。
本發明的藥物組合物可以為固體或液體的形式。一方面,載體是微粒,使得組合物例如為片劑或粉末的形式。載體可以是液體,所述組合物例如為口腔油、可注射的液體或氣霧劑(其例如可用于吸入給藥)。
在某些實施方案中,將治療化合物作為加壓氣霧劑或霧狀制劑通過吸入直接給予患者的肺。這類制劑可以含有多種已知的氣霧劑推進劑中的任何一種,所述氣霧劑推進劑可用于肺內和/或鼻內吸入給藥。此外,可以存在水,具有或不具有多種助溶劑、表面活性劑、穩定劑(例如,抗氧化劑、螯合劑、惰性氣體和緩沖液)中的任何一種。對于待給予的來自多劑量容器的組合物而言,通常添加抗微生物劑。這類組合物還通常被過濾和滅菌,并且可以被凍干以提供增強的穩定性和改善溶解度。
當意圖用于口服給藥時,藥物組合物優選為固體或液體形式,其中本文所認為的固體或液體形式包括半固體、半液體和凝膠形式。
作為用于口服給藥的固體組合物,可以將藥物組合物配制成粉末、顆粒、壓縮片劑、丸劑、膠囊、口香糖、薄脆餅或類似形式。這類固體組合物通常含有一種或多種惰性稀釋劑或可食用的載體。此外,可以存在一種或多種以下試劑:粘合劑,例如羧甲基纖維素、乙基纖維素、微晶纖維素、黃蓍膠或明膠;諸如淀粉、乳糖或糊精的賦形劑,諸如海藻酸、海藻酸鈉、primogel、玉米淀粉等的崩解劑;諸如硬脂酸鎂或sterotex的潤滑劑;諸如膠態二氧化硅的助流劑;諸如蔗糖或糖精的甜味劑;諸如薄荷、甲基水楊酸或橙香精的調味劑和著色劑。
當藥物組合物為膠囊,例如,明膠膠囊的形式時,除了上述類型的材料之外,其還可以含有諸如聚乙二醇或油的液體載體。
藥物組合物可以為液體形式,例如,酏劑、糖漿劑、溶液、乳劑或懸浮液。作為兩個實例,液體可用于口服給藥或用于注射遞送。當意圖用于口服給藥時,優選的組合物除了含有本化合物之外,還含有甜味劑、防腐劑、染料/著色劑和鮮味劑中的一種或多種。在意圖通過注射給藥的組合物中,可以含有表面活性劑、防腐劑、潤濕劑、分散劑、懸浮劑、緩沖液、穩定劑和等滲劑中的一種或多種。
本發明的液體藥物組合物,無論它們是溶液、懸浮液還是其它類似形式,都可以含有以下佐劑中的一種或多種:諸如注射用水的無菌稀釋液、鹽水、優選生理鹽水、林格氏液、等滲氯化鈉、諸如合成的甘油一酯或甘油二脂的固定油(可以充當溶劑或懸浮介質)、聚乙二醇、甘油、丙二醇或其它溶劑;諸如苯甲醇或對羥基苯甲酸甲酯的抗菌劑;諸如抗壞血酸或亞硫酸氫鈉的抗氧化劑;諸如乙二胺四乙酸的螯合劑;諸如醋酸鹽、檸檬酸鹽或磷酸鹽的緩沖液和諸如氯化鈉和右旋糖的用于調整滲透壓的試劑。可以將腸胃外制劑封裝在由玻璃或塑料制成的安瓿、一次性注射器或多劑量小瓶中。生理鹽水是優選的佐劑。可注射的藥物組合物優選是無菌的。
意圖用于腸胃外或口服給藥的本發明的液體藥物組合物含有本發明化合物的量應當使得可以獲取合適的劑量。通常,組合物中本發明化合物的量為至少0.01%。當意圖用于口服給藥時,該量可以在組合物重量的0.1-約70%間變化。某些口服藥物組合物含有約4%-約75%的本發明的化合物。按照本發明,某些藥物組合物和制劑被制備成一腸胃外劑量單位含有0.01-10%重量比的稀釋前的本發明的化合物。
本發明的藥物組合物可意圖用于局部給藥,在這種情況下,載體可適于包含溶液、乳劑、軟膏或凝膠基質,所述基質例如可以包含以下的一種或多種:凡士林、羊毛脂、聚乙二醇、蜂蠟、礦物油、諸如水和酒精的稀釋液以及乳化劑和穩定劑。用于局部給藥的藥物組合物中可以存在增稠劑。如果意圖用于經皮給藥,則所述組合物可以含有透皮貼劑或離子電滲療法裝置。局部制劑含有本發明化合物的濃度可以為約0.1-約10%w/v(每單位體積的重量)。
本發明的藥物組合物可意圖用于直腸給藥,例如為栓劑的形式,其可以在直腸中溶解并釋放藥物。用于直腸給藥的組合物可以含有油脂性基質作為合適的無刺激性賦形劑。這類基質包括但不限于羊毛脂、可可脂和聚乙二醇。
本發明的藥物組合物可以含有多種物質,其調整固體或液體劑量單位的物質形式。例如,組合物可以含有能在活性成分周圍形成包被外殼的材料。能形成包被外殼的材料通常是惰性的,并且可以選自例如,糖、蟲膠和其它腸衣包被劑。可選擇地,可以將活性成分包封在明膠膠囊中。
固體或液體形式的本發明的藥物組合物可以含有能與本發明化合物結合并進而促進所述化合物的遞送的試劑。具有這種能力的合適的試劑包括單克隆或多克隆抗體、蛋白或脂質體。
本發明的藥物組合物可以由可作為氣霧劑給藥的劑量單位組成。術語氣霧劑用來表示從膠體性質的系統到由加壓包裝組成的系統的多種系統。可以通過液化或壓縮的氣體或通過能分散活性成分的合適的泵送系統來實現遞送。為了遞送活性成分,本發明化合物的氣霧劑可以以單相、雙相或三相系統實現遞送。氣霧劑的遞送包括必需的容器、激活劑、泵、子容器等,它們在一起可以組成試劑盒。無需大量的實驗,本領域技術人員就可以確定優選的氣霧劑。
可以通過制藥領域內公知的方法來制備本發明的藥物組合物。例如,可以通過將本發明的化合物與無菌蒸餾水組合以便形成溶液來制備意圖通過注射給藥的藥物組合物。可以添加表面活性劑以促進均勻溶液或懸浮液的形成。表面活性劑是與本發明化合物非共價相互作用的化合物,以便促進化合物在水性遞送系統中的溶解或均勻懸浮。
本發明的化合物或其藥學可接受的鹽以治療有效量被給藥,該量隨多種因素變化,包括所用的具體化合物的活性、化合物的代謝穩定性和作用長度、患者的年齡、體重、一般健康、性別和飲食、給藥的方式和時間、排泄速度、藥物組成、具體病癥或疾病狀態的嚴重程度以及個體經歷的治療。通常,治療有效的日劑量(對于70kg的哺乳動物而言)為約0.001mg/kg(即0.07mg)至約100mg/kg(即7.0g);優選地,治療有效劑量(對于70kg的哺乳動物而言)為約0.01mg/kg(即0.7mg)至約50mg/kg(即3.5g);更優選地,治療有效劑量(對于70kg的哺乳動物而言)為約1mg/kg(即70mg)至約25mg/kg(即1.75g)。
在某些實施方案中,口服給予colv的劑量為每天.001mg至500mg。在一個具體的實施方案中,本文所述的colv的口服劑量為每天0.01mg至50mg。在另一實施方案中,本文所述的colv的口服劑量為每天0.1mg至0.5mg。在一個實施方案中,colv的口服劑量為每天0.01、0.02、0.03、0.04、0.05、0.06、0.07、0.08、0.09、or0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2.0、2.1、2.2、2.3、2.4、2.5、2.6、2.7、2.8、2.9、3.0、3.1、3.2、3.3、3.4、3.5、3.6、3.7、3.8、3.9、4.0、4.1、4.2、4.3、4.4、4.5、4.6、4.7、4.8、4.9或5.0mg。在另一實施方案中,colv的口服劑量可以為每天5.1、5.2、5.3、5.4、5.5、5.6、5.7、5.8、5.9、6.0、6.1、6.2、6.3、6.4、6.5、6.6、6.7、6.8、6.9、7.0、7.1、7.2、7.3、7.4、7.5、7.6、7.7、7.8、7.9或8.0mg。在某些實施方案中,劑量可以單一劑量給予或可以在一天內分多劑量給予,例如每天2、3或4次劑量,總共為每天具體的mg劑量。
如本文其它地方所描述,在某些實施方案中,本文所用的治療有效劑量的colv是利用本文所述的各種方法檢測到的足以誘導對colv的耐受性的劑量。在某些實施方案中,如通過本文所述的方法例如elisa所檢測的,對colv的耐受性的誘導導致血清抗colv抗體減少。在另一實施方案中,本文所用的治療有效劑量的colv是利用本文所述的各種方法檢測到的足以誘導t細胞對colv的耐受性的劑量,所述方法例如細胞因子釋放測定、細胞內細胞因子染色和流式細胞術、elispot等。還可以利用功能性t細胞測定,例如細胞毒性增殖測定。
本發明的化合物或其藥學可接受的鹽可以與一種或多種其它治療劑同時給藥,在其之前給藥或在其之后給藥。這種組合療法包括單一藥物劑量制劑的給藥以及本發明化合物的給藥和每種活性劑在其自身單獨的藥物劑量制劑中,所述單一藥物劑量制劑含有本發明的化合物和一種或多種其它活性劑。例如,本發明的化合物和另一種活性劑可以在單一的口服劑量組合物(例如片劑或膠囊)中一起給予患者,或者每種試劑以單獨的口服劑量制劑給藥。如果使用單獨的劑量制劑,本發明的化合物和一種或多種其它活性劑可以基本上在同一時間即同時給藥,或者以單獨的錯開的時間即相繼給藥;組合療法應被理解為包括所有這些方案。
本發明的化合物可以給予遭受本文所述疾病或病癥折磨的個體,例如copd和重度且持續的哮喘。對于體內治療人類疾病而言,通常在給藥之前,將本文所述的化合物并入藥物組合物中。藥物組合物包含一種或多種本文所述的化合物和本文其它地方所述的藥學可接受的載體或賦形劑。為了制備藥物組合物,將有效量的一種或多種化合物與本領域技術人員已知的適用于特定給藥方式的任何藥物載體或賦形劑混合。藥物載體可以是液體、半固體或固體。用于腸胃外、皮內、皮下或局部應用的溶液或懸浮液可以包括例如,無菌稀釋液(例如水)、鹽水、固定油、聚乙二醇、甘油、丙二醇或其它合成的溶劑;抗微生物劑(例如苯甲醇和對羥基苯甲酸甲酯);抗氧化劑(例如抗壞血酸和亞硫酸氫鈉)和螯合劑(例如乙二胺四乙酸(edta));緩沖液(例如醋酸鹽、檸檬酸鹽和磷酸鹽)。如果靜脈內給藥,合適的載體包括生理鹽水或磷酸鹽緩沖鹽水(pbs)和含有增稠劑和加溶劑(例如葡萄糖、聚乙二醇、聚丙二醇和以上的混合物)的溶液。
本文所述的化合物可以與保護其免于從身體快速排除的載體一起制備,例如時間釋放制劑或包被。這類載體包括受控釋放制劑,例如但不限于,植入物和裝入微膠囊的遞送系統,以及生物可降解的、生物兼容的聚合物,例如乙烯醋酸乙烯酯、聚酐、聚乙醇酸、聚原酸酯、聚乳酸和本領域技術人員已知的其它聚合物。
在某些實施方案中,能促進誘導耐受性的佐劑包括地塞米松(參見例如,y.kang,etal.,j.immunol.2008,180:5172-5176)、脂多糖(lps)和霍亂毒素亞基,并且可以被添加至制劑。還考慮將某些其它耐受原性載體與本發明的colv組合物一起使用。這類載體包括礦物油載體,例如弗氏不完全佐劑(ifa)或弗氏完全佐劑(cfa)。ifa是礦物油的乳劑。cfa是含有各種量的殺死的分支桿菌有機體的礦物油制劑。然而,ifa和cfa不允許用于人類,因為礦物油是不可代謝的,并且不能被人體降解。
在某些實施方案中,脂肪乳劑,其已用于人患者的靜脈營養很多年了,其還可以作為利用本發明多肽的耐受原性多肽療法的媒介。這類乳劑的兩個實例是可購買到的稱為intralipid和lipofundin的脂肪乳劑。“intralipid”是kabipharmacia,sweden的注冊商標,是用于靜脈營養的脂肪乳劑,描述于美國專利第3,169,094號中。“lipofundin”是b.braunmelsungen,germany的注冊商標。兩種均含有諸如脂肪的大豆油(1,000ml蒸餾水中100或200g:分別為10%或20%)。蛋黃磷脂是intralipid中所用的乳化劑(12g/l蒸餾水),且lipofundin中是蛋黃卵磷脂(12g/l蒸餾水)。intralipid和lipofundin兩者中的等滲性均由添加甘油(25g/l)導致。這些媒介被認為是實際的生物活性載體,當其與可疑自體抗原混合時,促進自身免疫t細胞的th1至th2的轉移。在某些實施方案中,這類媒介是這樣的脂肪乳劑,其含有10-20%的植物和/或動物來源的甘油三磷脂、1.2-2.4%的植物和/或動物來源的磷脂、2.25-4.5%滲透壓調節劑、0-0.05%抗氧化劑并用無菌水加至100%。
在某些實施方案中,colv或其耐受原性片段可以連接至白喉毒素受體以增強gi攝入。
本發明的耐受原性組合物可用于治療哮喘和copd和colv自身免疫相關的其它肺病。因此,在一個實施方案中,本發明提供了用于治療慢性阻塞性肺病的方法,包括向所述copd患者給予治療有效量的v型膠原、或其耐受原性片段或免疫原性片段,所述免疫原性片段以有效誘導耐受性的劑量給予。在這方面中,copd患者可以患肺氣腫或慢性阻塞性支氣管炎。
本發明另一方面提供了用于治療哮喘的方法,包括向所述哮喘患者給予治療有效量的v型膠原或其耐受原性片段或免疫原性片段,所述免疫原性片段以有效誘導耐受性的劑量給予。在這方面中,哮喘患者可以患有重度、持續的哮喘。在另一實施方案中,本發明提供了用于降低哮喘患者哮喘嚴重程度的方法。
本發明還提供了用于預防有發展成慢性阻塞性肺病風險的個體中慢性阻塞性肺病的發展的方法,包括向所述個體給予治療有效量的v型膠原或其耐受原性片段。
如本文其它地方所指出的,可以給予完整的colv或其成分α鏈的任何一種或多種或上述任一分子的耐受原性片段。此外,同樣如本文其它地方所指出的,可以按本文所述的任何方法,使用有效誘導對colv的耐受性的劑量,給予完整的colv或其成分α鏈的任何一種或多種的免疫原性片段。這種劑量可以隨多種因素變化,包括所用的具體蛋白或片段的活性、這些化合物的代謝穩定性和作用長度、患者的年齡、體重、一般健康、性別和飲食、給藥的方式和時間、排泄速度、藥物組成、具體病癥或疾病狀態的嚴重程度以及個體經歷的治療。因此,這種耐受原性劑量可以使用已知的測驗方案通過經驗來確定或者通過在領域內已知的模型系統中測驗組合物并外推來確定。還可以進行可控的臨床試驗。劑量還可以隨待緩解的疾病狀態的嚴重程度而改變。
如同本領域技術人員所理解的,多種因素可被評價從而確定用于治療、預防或降低copd或哮喘的嚴重程度的本發明的化合物和方法的有效性。這些因素包括可為臨床醫生進行評價的哮喘或copd的典型臨床癥狀。這些癥狀可以包括但不限于:對于哮喘而言:哮鳴、胸部緊迫感或疼痛、心率加快、發汗、如通過最大流量計所測量的流速最高、咳嗽頻繁,尤其在夜間、容易喘不上氣或氣短、當運動時感到非常疲憊或虛弱、運動后哮鳴或咳嗽、感到疲憊、易煩躁、壞脾氣、或情緒多變、如通過最大流量計所測量的肺功能減弱或改變、傷寒或過敏癥(打噴嚏、流鼻涕、咳嗽、鼻塞、咽喉痛和頭痛)的跡象、失眠。這些因素包括copd的典型臨床癥狀,例如但不限于,產生的痰的量、痰的密度和粘性、痰的顏色和痰中帶血、氣短的嚴重程度、咳嗽和/或哮鳴;腳踝腫脹;健忘、糊涂、言語含糊和瞌睡;失眠;使用更多的枕頭或睡在椅子上而不是床上,以避免氣短;體重莫名其妙的增加或降低;持續性疲勞和沒勁;喪失性沖動;早晨頭痛、頭暈目眩、心神不定。
本發明的組合物和方法可以與用于copd和哮喘的其它已知治療聯合使用,例如但不限于皮質類固醇(例如,強的松、氟替卡松、甲基強的松龍)、支氣管擴張藥(例如,短效和長效的β2-激動劑、茶堿、吡布特羅、麻黃堿、舒喘靈、沙美特羅、左沙丁胺醇、雙氯醇胺異丙托溴胺)和白三烯調節劑(例如,孟魯司特、扎魯司特、棄白通)。
參考文獻
以下參考文獻通過引用具體并入本文,它們提供示例性方案或其它詳情的程度補充了本文所示的那些實施方案。chen,inobe,marks,gonnella,kuchroo,weiner,“peripheraldeletionofantigen-reactivetcellsinoraltolerance(口服耐受性中抗原反應性t細胞的外部刪除)”nature,376:177-180,1995.chen,kuchroo,inobe,hafler,weiner,"regulatoryt-cellclonesinducedbyoraltolerance:suppressionofautoimmuneencephalomyelitis(由口服耐受性誘導的調節型t細胞克隆:自身免疫性腦脊髓炎的抑制)"science,265:1237-1240,1994.chiang,mainardi,seyer,"typev(a-b)collageninducesplateletaggregation(v(a-b)型膠原誘導血小板聚集)"j.lab.clin.med.,95:99-107,1980.cremer,ye,terato,owens,seyer,kang,"typexicollagen-inducedarthritisinthelewisrat:characterizationofcellularandhumoralimmuneresponsestonativetypesxi,v,andiicollagenandconstituentα-chains(路易鼠中xi型膠原誘導的關節炎:對天然的xi型、v型和ii型膠原和成分α鏈的細胞和體液免疫應答的表征)"j.immunol.153:824-832,1994.danzer,kirchner,rink,"cytokineinteractionsinhumanmixedlymphocyteculture(人類混合淋巴細胞混合物中細胞因子的相互作用)"transplantation,57(11):1638-1642,1994.demeester,rolfe,kunkel,swiderski,lincoln,deeb,strieter,"thebimodalexpressionoftumornecrosisfactor-αinassociationwithratlungreimplantationandallograftrejection(腫瘤壞死因子α的雙峰表達與大鼠肺再植入和同種移植物排斥相關)"j.immunol.,150(6):2494-2505,1993.fariaandweiner,"oraltolerance:mechanismsandtherapeuticapplications(口服耐受性:機制和治療應用)"adv.immunol.,73:153-264,1999.fedoseyeva,zhang,orr,levin,buncke,benichou,"denovoautoimmunitytocardiacmyosinafterhearttransplantationanditscontributiontotherejectionprocess(心臟移植后對心肌肌球蛋白的原位自身免疫及其對排斥過程的貢獻)"j.immunol.,162:6836-42,1999.garrovillo,ali,oluwole,"indirectallorecognitioninacquiredthymictolerance:inductionofdonor-specifictolerancetoratcardiacallograftsbyallopeptide-pulsedhostdendriticcells(獲取的胸腺耐受性中間接的同種識別:由同種肽脈沖的宿主樹突細胞誘導對大鼠心臟同種移植物的供體特異性耐受)"transplantation,68:1827-1834,1999.hancock,sayegh,kwok,weiner,carpenter,"oral,butnotintravenous,alloantigenpreventsacceleratedallograftrejectionbyselectiveintragraftth2cellactivation(口服而非靜脈內的同種抗原通過選擇性內移植物th2細胞激活防止加速的同種移植物排斥)"transplantation,55:1112-1118,1993.hanson,gorman,oui,cheah,solomon,trowsdale,"thehuman2(xi)collagengene(col11a2)mapstothecentromericofthemajorhistocompatibilitycomplexonchromosome6(人類α2(xi)膠原基因(col11a2)繪制在6號染色體的主要組織相容性復合體的著絲粒上)"genomics,5:925-931,1989.hirt,you,moller,boeke,starke,spranger,wottge,"developmentofobliterativebronchiolitisafterallogeneicratlungtransplantation:implicationofacuterejectionandthetimepointoftreatment(同種大鼠肺移植后閉塞性細支氣管炎的發展:涉及急性排斥和治療的時間點)"j.heartlungtransplant.,18:542-548,1999.huang,fuchimoto,scheier-dolberg,murphy,neville,sachs,j.clin.invest.,105:173-181,2000.ishido,matsuoka,matsuno,nakagawa,tanaka,"inductionofdonor-specifichyporesponsivenessandprolongationofcardiacallograftsurvivalbyjejunaladministrationofdonorsplenocytes(由供體脾細胞的空腸給藥所誘導的供體特異的低反應性和心臟同種移植物存活的延長)"transplantation,68:1377-1382,1999.iyer,woo,cornejo,gao,mccoubrey,maines,buelow,"characterizationandbiologicsignificanceofimmunosuppressivepeptided2702.75-84(eαv)bindingprotein(免疫抑制肽d2702.75-84(eαv)結合蛋白的表征和生物學意義)"j.bio.chem.,273(5):2692-2697,1998.joo,pepose,stuart,"t-cellmediatedresponsesinamurinemodeloforthotopiccornealtransplantation(常位角膜移植的鼠模型中t細胞介導的應答)"invest.ophthalmol.vis.sci.,36:1530-1540,1995.konomi,hayashi,nakayasu,arima,"localizationoftypevcollagenandtypeivcollageninhumancornea,lung,andskin(人類角膜、肺和皮膚中v型膠原和iv型膠原的定位)"am.j.pathol.,116:417-426,1984.krenskyandclayberger,"hla-derivedpeptidesasnovelimmunosuppressives(hla來源的肽作為新的免疫抑制劑)"nephrol.dial.transplant.,12:865-878,1997.lowry,marghesco,blackburn,"immunemechanismsinorganallograftrejection.vi.delayed-typehypersensitivityandlymphotoxininexperimentalrenalallograftrejection(器官同種移植排斥的免疫機制,vi.實驗腎臟同種移植排斥中的延遲型超敏性和淋巴毒素)"transplantation.,40:183-188,1985.madriandfurthmayr,"collagenpolymorphisminthelung(肺中的膠原多態性)"humanpathology,11:353-366,1980.madriandfurthmayr,"isolationandtissuelocalizationoftypeab2collagenfromnormallungparenchyma(來自正常肺實質的ab2型膠原的分離和組織定位)"am.j.pathol.,94:323-332,1979.marck,prop,widevuur,"lungtransplantationintherat.iii.functionalstudiesiniso-andallografts(大鼠中的肺移植,iii.同體移植和同種移植的功能研究)"j.surgicalres.,35:149-158,1983.matsumura,marchevsky,zuo,kass,matloff,jordan,"assessmentofpathologicalchangesassociatedwithchronicallograftrejectionandtoleranceintwoexperimentalmodelsofratlungtransplantation(在大鼠肺移植的兩個實驗模型中對與慢性同種移植排斥相關的病理學改變的評價)"transplantation.,59:1509-1517,1995.morrisandbachinger,"typexicollagenisaheterotrimerwiththecomposition(1α,2α,3α)retainingnon-triplehelicaldomains(xi型膠原是保留非三螺旋結構域的組分(1α、2α、3α)的異三聚體).j.biologicalchem.,262:11345-11350,1987.murphy,magee,alexander,waaga,snoeck,vella,carpenter,sayagh,"inhibitionofallorecognitionbyahumanclassiimhc-derivedpeptidethroughtheinductionofapoptosis(人ii類mhc來源的肽通過誘導凋亡抑制同種識別)"j.clin.invest.,103:859-867,1999.nosner,goldberg,naftzger,lyu,clayberger,krensky,"hla-derivedpeptideswhichinhibittcellfunctionbindtomembersoftheheat-shockprotein70family(抑制t細胞功能的hla來源的肽與熱激蛋白70家族的成員結合)"j.exp.med.,183:339-348,1996.oluwole,chowdhury,jin,hardy,"inductionoftransplantationintolerancetoratcardiacallograftsbyintrathymicinoculationofallogeneicsolublepeptides(通過同種可溶肽的胸腺內接種移植誘導對大鼠心臟同種移植物的不耐受)"transplantation,56(6):1523-1527,1993.prop,nieuwenhuis,wildevuur,"lungallograftrejectionintherat.i.acceleratedrejectioncausedbygraftlymphocytes(大鼠中的肺同種移植排斥,i.移植淋巴細胞導致加速排斥)"transplantation,40:25-30,1985.prop,wildevuur,nieuwenhuis,"lungallograftrejectionintherat.ii.specificimmunologicalpropertiesoflunggrafts(大鼠中的肺同種移植排斥,ii.肺移植的特異性免疫學特性)"transplantation,40:126-131,1985.sayagh,watschinger,carpenter,"mechanismsoftcellrecognitionofalloantigen(同種抗原的t細胞識別的機制)"transplantation,57:(9)1295-1302,1994.sayeghandkrensky,"novelimmunotherapeuticstrategiesusingmhcderivedpeptides(利用mhc來源的肽的新的免疫治療策略)"kidneyint.suppl.53:s13-20,1996.sayegh,khoury,hancock,weiner,carpenter,"inductionofimmunityandoraltolerancewithpolymorphicclassiimajorhistocompatibilitycomplexallopeptidesintherat(利用多態ii類主要組織相容性復合體同種肽在大鼠中誘導的免疫性和口服耐受)"proc.natl.acad.sci.,89:7762-7766,1992.sayegh,zhang,hancock,kwok,carpenter,weiner,"down-regulationoftheimmuneresponsetohistocompatibilityantigensandpreventionofsensitizationbyskinallograftsbyorallyadministeredalloantigen(通過口服給予同種抗原下調對組織相容性抗原的免疫應答和防止皮膚同種移植物的致敏作用)"transplantation,53:163-166,1992.sekine,nowen,heidler,vanrooijen,brown,cummings,wilkes,"roleofpassengerleukocytesinallograftrejection--effectofdepletiondonoralveolarmacrophagesonthelocalproductionoftnf-alpha,thelper1/thelper2cytokines,iggsubclasses,andpathologyinaratmodeloflungtransplantation(過客白細胞在同種移植排斥中的作用--耗盡供體肺泡巨噬細胞對肺移植的大鼠模型中tnf-α、t輔助1/t輔助2細胞因子、igg亞類的局部產生以及病理的影響)"j.immunol,159:4084-4093,1997.seyerandkang,"covalentstructureofcollagen:aminoacidsequenceofthreecyanogenbromide-derivedpeptidesfromhumanalpha1(v)collagenchain(膠原的共價結構:來自人α1(v)膠原鏈的三種溴化氰衍生的肽的氨基酸序列).arch.biochem.biophys.271(1):120-129,1989.sivasai,smith,poindexter,sundaresan,trulock,lynch,cooper,patterson,mohanakumar,"indirectrecognitionofdonorhlaclassipeptidesinlungtransplantrecipientswithbronchiolitisobliteranssyndrome(在患閉塞性細支氣管炎綜合征的肺移植接受者中供體hlai類肽的間接識別)"transplantation.67(8):1094-1098,1999.smithjr,williams,brandt,"interactionofproteoglycanswithpericellular(1alpha,2alpha,3alpha)collagensofcartilage(蛋白聚糖與軟骨細胞周圍(1α、2α、3α)膠原的相互作用)"j.biol.chem.,260:10761-10767,1985.starkandostrow,trainingmanualseries,laboratoryanimaltechnician,americanassociationforlaboratoryanimalscience,181-182,1990.stroberandcoffman,"toleranceandimmunityinthemucosalimmunesystem(粘膜免疫系統中的耐受性和免疫性)"res.immunol.,148:489-599,1997.trulock,"lungtransplantation(肺移植)"am.j.respir.crit.caremed.,155:789-818,1997.vanbuskirk,wakely,sirak,orosz,"patternsofallosensitizationinallograftrecipients:long-termallograftacceptanceisassociatedwithactivealloantibodyproductioninconjunctionwithactiveinhibitionofalloreactivedelayed-typehypersensitivity(同種移植接受者中的同種致敏模式:長期同種移植接受與活性同種抗體產生和同種反應性遲發型超敏反應的活性抑制相關)"transplantation.,65:1115-1123,1998.westra,prop,kuijpers,"aparadoxinheartandlungrejection(心臟和肺排斥中的悖論)"transplantation,49:826-828,1990.whitacre,gienapp,orosz,bitar,"oraltoleranceinexperimentalautoimmuneencephalomyelitis.iii.evidenceforclonalanergy(實驗自身免疫性腦脊髓炎中的口服耐受,iii.克隆無反應的證據)"j.immunol.,147:2155-2163,1991.wilkes,bowman,cummings,heidler,"allogeneicbronchoalveolarlavagecellsinducethehistologyandimmunologyoflungallograftrejectioninrecipientmurinelungs.roleoficam-1ondonorcell(異源支氣管肺泡灌洗液細胞誘導接受鼠的肺中肺同種移植排斥的組織學和免疫學.供體細胞上icam-1的作用)"transplantation,67(6):890-896,1999.wilkes,heidler,bowen,quinlan,doyle,cummings,doerschuk,"allogeneicbronchoalveolarlavagecellsinducethehistologyofacutelungallograftrejection,anddepositionofigg2ainrecipientmurinelungs(異源支氣管肺泡灌洗液細胞誘導急性同種移植排斥的組織學以及igg2a接受鼠的肺中的沉積)"j.immunol.,155:2775-2783,1995.wilkes,thompson,cummings,bragg,heidler,"instillationofallogeneiclungmacrophagesanddendriticcellscausedifferentialeffectsonlocalify-γproduction,lymphocyticbronchitis,andvasculitisinrecipientmurinelungs(異源肺巨噬細胞和樹突細胞灌輸對接受鼠的肺中局部ify-γ產生、淋巴細胞性支氣管炎和脈管炎產生不同的影響)"j.leukoc.biol.64:578-586,1998.wilson,ebringer,ahmadi,wrigglesworth,tiwana,fielder,binder,ettelaie,cunningham,joannou,bansal,""sharedaminoacidsequencesbetweenmajorhistocompatibilitycomplexclassiiglycoproteins,typexicollagenandproteusmirabilisinrheumatoidarthritis(主要組織相容性復合體ii類糖蛋白、xi型膠原和類風濕性關節炎中奇異變形桿菌之間共有的氨基酸序列)"ann.rheum.dis.,"54:216-220,1995.woessnerjr.,"thedeterminationofhydroxyprolineintissueandproteinsamplescontainingsmallproportionsofthisimminoacid(對含有少量羥脯氨酸的組織和蛋白樣品中該氨基酸的測定)"arch.biochem.biophys.93:440-447,1961.yagyu,steinhoff,schafers,dammenhayn,haverich,borst,"comparisonofmononuclearcellpopulationsinbrochoalveolarlavagefluidinacuterejectionafterlungtransplantationandmycoplasmainfectioninrats(對肺移植后急性排斥和大鼠支原體感染的支氣管肺泡灌洗液中單核細胞群體的比較)"j.hearttransplant.,9:516-525,1990.yamagami,tsuru,ohkawa,endo,isobe,"suppressionofallograftrejectionwithanti-alphabetatcellreceptorantibodyinratcornealtransplantation(在大鼠角膜移植中用抗αβt細胞受體的抗體抑制同種移植排斥)"transplantation,67:600-604,1999.yoshino,quattrocchi,weiner,"suppressionofantigen-inducedarthritisinlewisratsbyoraladministrationoftypeiicollagen(在路易鼠中通過口服給予ii型膠原抑制抗原誘導的關節炎)"arthritisrheum.38:1092-1096,1995.yousem,berry,cagle,chamberlain,husain,hruban,marchevsky,ohori,ritter,stewart,tazelaar,""revisionofthe1990workingformulationfortheclassificationofpulmonaryallograftrejection:lungrejectionstudygroup(對1990年從事肺同種移植排斥分類的概述:肺排斥研究小組)"j.heartlungtransplant,15:1-15,1996.zheng,markees,hancock,li,greine,li,mordes,sayegh,rossini,strom,"ctla4signalsarerequiredtooptimallyinduceallografttolerancewithcombineddonor-specifictransfusionandanti-cd154monoclonalantibodytreatment(需要ctla4信號聯合供體特異的灌輸和抗cd154單克隆抗體治療來優化誘導同種移植耐受)"j.immunol.,162:4983-4990,1999.
實施例
實施例1
copd患者中抗v型膠原抗體增加
血漿獲自正常的志愿者(不吸煙的成年人,年齡為18-55)和被證實患有肺氣腫的志愿者。按照wo2007/120947中所描述的流式細胞術珠子測定法檢測copd患者和對照健康個體中抗colv抗體的水平。
簡單而言,1)用無菌pbs將鏈霉親和素包被的珠子(5μm,結合能力:10-20μg/1×107珠子(polyscience,warrington,pa))洗滌兩次。將珠子(1×107)懸浮于100μl具有40μg人v型膠原的pbs中,并于4℃孵育60分鐘。2)用20μm兔抗人v型膠原抗體的抗體(bioten)(abeam,cambridge,ma),按照上述1中相同的方法產生陽性對照。3)對于每次測定,將1×106偶聯的珠子在pbs中洗滌兩次,并在加50μl血清的100μlpbs中進行孵育。室溫下孵育30分鐘后,用含有10%fcs的pbs將珠子洗滌三次。4)將珠子懸浮于無菌pbs+10%fbs中,并在室溫下與二抗孵育大約30分鐘。通常,使用與r-pe偶聯的5μl抗人igg抗體(sigma,saintlouis)。將珠子在含有10%fcs的pbs中洗滌三次,懸浮于300μlpbs/fcs溶液中,并利用流式細胞儀進行分析。
如圖1所示,與對照相比(n=42),copd患者(n=16)中抗v型膠原抗體顯著升高。
實施例2
哮喘患者中抗v型膠原抗體增加
血漿獲自正常的志愿者(不吸煙的成年人,年齡為18-55)和被證實患有慢性哮喘的志愿者。用實施例1中概括的珠子測定法檢測哮喘患者和對照志愿者個體中抗colv抗體的水平。
如圖2所示,在20名哮喘患者中發現8名患者的抗v型膠原抗體的水平提高。
實施例3
小鼠靜脈內的v型膠原防止卵白蛋白誘導的氣道高反應性
本實施例證實,在該建立良好的鼠哮喘模型中,v型膠原的靜脈內給藥防止卵白蛋白誘導的氣道高反應性。
用100μg單獨的col(v)、或混在弗氏完全佐劑(cfa)中的col(v)、或pbs或單獨的cfa經尾靜脈注射balb/c小鼠。7天之后,小鼠接受于明礬中的卵白蛋白的ip注射,7天之后重復一次。在最后一次卵白蛋白/明礬注射之后7天,用增加劑量的氣霧化卵白蛋白激發每組中的小鼠,隨后測量氣道阻力(penh)。
結果顯示,單獨的col(v)消除了卵白蛋白誘導的氣道高反應性(參見圖3)。進行其它的實驗來證實這些結果。這些結果總結在以下的表2中。
實施例4
靜脈內的col(v)在肺單核細胞中誘導ifn-γ轉錄本
本實施例顯示,單獨col(v)的靜脈注射在肺單核細胞中誘導th1應答,特征為誘導出ifn-γ轉錄本。用100μg單獨的col(v)經尾靜脈、單獨的cfa在尾基部、colv加cfa在尾基部或單獨的pbs經靜脈對balb/c小鼠進行注射。7天之后,小鼠接受于明礬中的卵白蛋白的ip注射,7天之后重復一次。在最后一次卵白蛋白/明礬注射之后7天,用增加劑量的氣霧化乙酰甲膽堿激發每組中的小鼠,隨后測量響應于乙酰甲膽堿激發的氣道阻力(penh)。從每個處理組的小鼠的肺實質分離的單核細胞中提取rna,并進行定量pcr用于測定il-4、il-5、il-13、ifn-γ和il-10的表達水平(參見圖4;數據代表從每組5只小鼠的肺單核細胞合并的rna)。
實驗結果指示,靜脈內單獨給予col(v)在肺單核細胞中誘導出ifn-γ轉錄本。ifn-γ拮抗il-13和il4,il-13和il4兩種細胞因子被認為在哮喘發病機理中起重要作用。因此,不希望受理論的束縛,對抗il4/il13效應的該th1應答的誘導在卵白蛋白誘導的哮喘中colv介導的保護性效應中起作用。
本說明書中涉及的和/或申請數據表中列舉的所有上述美國專利、美國專利申請公開、美國專利申請、外國專利、外國專利申請和非專利出版物都通過引用全文并入本文。
從上述應當理解,盡管為了說明的目的,本文描述了本發明的具體實施方案,但是在不脫離本發明的實質和范圍的條件下可以進行多種修改。因此,本發明僅受所附權利要求書的限制。
序列表
<110>印第安那大學科技研究公司
<120>用于治療慢性阻塞性肺病和哮喘的組合物和方法
<130>480266.404pc
<140>pct
<141>2010-04-22
<150>us61/266,048
<151>2009-12-02
<150>us61/171,705
<151>2009-04-22
<160>6
<170>fastseqforwindowsversion4.0
<210>1
<211>8439
<212>dna
<213>智人
<400>1
cgcactctccgtccccgcggctggcgcaggacctcactcgagcggagcgcccacggggag60
cgggtcgcggggcggcggcggcgaggaggaggcgagaaggagttggaggaggaggaggag120
gaggcgagggcgagctagcccagcggggtcccggccgccccgcgggccaaagtcgagccc180
tcccgcccgtgggcgagcgcgccagccgccccttccagaacagccgccgccacaaagaag240
aacggggggtgccgaggtccccatgacctcctaaagtggtgcggtccctgctgagtgcgc300
tgcccgggccgtgacccgcgcccctgtgcgtccccgcgcgcctccgagcgcccctgtgcg360
ccccggcccgcgccccgccggcatggacgtccatacccgctggaaagcgcgcagcgcgct420
ccgcccgggcgccccgctgctgcccccgctgctgctgctgctgctgtgggcgccgcctcc480
gagccgcgcagctcagccagcagatctcctgaaggttctagattttcacaacttgcctga540
tggaataacaaagacaacaggcttttgcgccacgcggcgatcttccaaaggcccggatgt600
cgcttacagagtcaccaaagacgcgcagctcagcgcacccaccaagcagctgtaccctgc660
gtctgcatttcccgaggacttctccatcctaacaactgtgaaagccaagaaaggcagcca720
ggccttcctggtctccatctacaacgagcagggtatccagcagattgggctggagctggg780
ccgctctcccgtcttcctctacgaggaccacacggggaagcctggcccggaagactaccc840
cctcttccggggcatcaacctgtcagatggcaagtggcacagaattgctctcagcgtcca900
caagaaaaatgtcaccttgatcctcgactgtaaaaagaagaccaccaaattcctcgaccg960
cagcgaccaccccatgatcgacatcaatggcatcatcgtgtttggcacccggatcctgga1020
tgaggaggtgtttgagggtgacatccagcagctgctctttgtctcggaccaccgggcagc1080
ttatgattactgtgagcactacagccctgactgtgacaccgcagtacctgacaccccaca1140
gtcgcaggaccccaatccagatgaatattacacggaaggagacggcgagggtgagaccta1200
ttactacgaatacccctactacgaagaccccgaagacctagggaaggagcccacccccag1260
caagaagcccgtggaagctgccaaagaaaccacagaggtccccgaggagctgaccccgac1320
ccccacggaagctgctcccatgcctgaaaccagtgaaggggctgggaaggaagaggacgt1380
cggcatcggggactatgactacgtgcccagtgaggactactacacgccctcaccgtatga1440
tgacctcacctatggcgagggggaggagaaccccgaccagcccacagacccaggcgctgg1500
ggccgaaattcccaccagcaccgccgacacctccaactcctccaatccagctccgcctcc1560
aggggaaggtgcggatgacttggagggggagttcactgaggaaacgatccggaaccttga1620
cgagaactactacgacccctactacgaccccaccagctccccgtcggagatcgggccggg1680
aatgccggcgaaccaggataccatctatgaagggattggaggacctcggggcgagaaagg1740
ccaaaagggagaaccagcgattatcgagccgggcatgctcatcgagggcccgcctggccc1800
agaaggccccgcgggtcttcccggacctccaggaaccatgggtcccactggccaagtcgg1860
ggaccctggagaaaggggcccccctggacgcccaggccttcctggggccgatggcctgcc1920
cggtcctccaggaaccatgctcatgctgcccttccggtttggaggtggcggcgatgcggg1980
ctccaaaggccccatggtctcagcccaggagtcccaggcgcaagccattctccagcaggc2040
caggttggcactgaggggaccagctggcccgatgggtctcacagggagacctggccctgt2100
gggtccccctgggagcggaggtttgaagggcgagccgggagacgtggggcctcagggtcc2160
tcgaggtgtgcaaggcccgcctggtccggccgggaagcccggaagacggggtcgggctgg2220
gagtgatggagccagaggaatgcctggacaaactggccccaagggtgaccggggtttcga2280
cggcctggctgggttgccaggcgagaagggccacaggggtgaccctggtccttccggccc2340
accaggacctccgggagacgatggagaaaggggtgacgacggagaagttgggcccagggg2400
gctgcctggggagcccgggccacgtggtctgcttgggccgaaggggcccccaggtcctcc2460
cggacctcccggtgtcacgggtatggacggccagccggggccaaaaggaaatgtgggtcc2520
ccagggagagcctggccccccaggacagcagggtaatccaggcgcccagggtcttccagg2580
cccccagggtgcaattggtcctccaggagaaaagggtcccttggggaaaccaggccttcc2640
aggaatgcccggtgctgacggacccccgggacaccctggcaaagaaggccctccaggaga2700
gaaaggaggtcagggtccacctggcccccagggtccgattggctacccaggtcctcgagg2760
agtcaagggggccgatggcatccgtggtctgaagggcacaaagggcgagaagggtgaaga2820
cggctttcctgggtttaaaggagacatgggcatcaagggtgatcggggggagatcggccc2880
acccggtcccaggggagaagatggccctgaaggcccaaagggtcgcggaggtcccaatgg2940
tgaccccggtcctctgggaccccctggggagaagggaaaactcggagtcccagggttacc3000
agggtatccaggaagacaaggaccaaagggctctattggattccctggatttcctggcgc3060
caatggagagaagggcggcagggggacccctggaaagccaggaccgcgggggcagcgagg3120
cccaacgggtccgaggggtgaaagaggcccccggggcatcactgggaagcctggccccaa3180
gggcaactccggaggtgacggcccagctggccctcctggtgaacggggacccaatggacc3240
ccaaggacccacaggatttcctggaccaaagggcccccctggccctccaggcaaggatgg3300
actcccaggacaccctggacagagaggcgagactggtttccaaggcaagaccggccctcc3360
aggcccccccggcgtggtcggccctcagggtcccacgggagaaacgggcccaatgggtga3420
gcgtggccaccctgggccccctggaccccccggtgaacaggggcttccgggccttgctgg3480
aaaagaagggacgaagggtgacccaggccctgcaggcctccctgggaaagatggccctcc3540
aggattacgtggtttccctggggaccgagggcttcctggtccagtgggagctcttggact3600
gaaaggcaatgaagggccccctggcccaccaggccctgcgggatctccaggggagagagg3660
tccagctggagccgctgggcccatcggaattccagggagacctgggccccagggaccccc3720
agggccggcaggagagaaaggggctcctggcgagaaaggcccacaaggcccagctggccg3780
agacggtctccaggggcctgtggggctcccgggtccagctggccctgtgggtccccctgg3840
agaagacggagataagggagagatcggggagccggggcagaaaggaagcaagggggacaa3900
aggagaacagggtcctcctgggcctacaggtcctcaaggccccatcggacagccaggccc3960
ctctggagctgacggcgagccggggcctcggggccagcagggccttttcgggcagaaagg4020
tgatgaaggtcccagaggctttcctggaccccctgggccagtggggctgcagggtttgcc4080
aggacctccaggcgagaagggtgagacaggagacgtgggccagatgggccccccgggtcc4140
ccctggcccccgaggaccctccggagctccaggtgctgatggcccacaaggtcccccagg4200
tggaataggaaaccctggtgcagtgggagagaagggcgagcctggcgaagcaggtgagcc4260
tggccttccgggagaaggcggccccccgggacccaaaggagaaaggggagagaagggcga4320
gtcaggcccttcaggtgctgccggaccccctggacccaaaggccctcccggagatgatgg4380
tcccaaaggcagccctggcccagtgggttttcctggagatcctggcccccccggagagcc4440
tggccccgcgggtcaagatggtccccctggtgacaaaggagatgatggtgaacccgggca4500
gacgggatcccccggccctactggtgaaccaggtccatcggggcctccaggaaaaagggg4560
tcccccaggccccgcaggccccgaaggcagacagggagagaaaggggccaagggagaagc4620
cggcttggaaggccctcctgggaagactggccccatcggcccccagggggcccctgggaa4680
gcccggaccggatggccttcgagggatccctggccctgtgggagaacaaggtctcccagg4740
atccccaggcccggacggtccccccggccccatgggtcccccaggacttcccggcctcaa4800
aggagattctggtcccaaaggtgaaaagggtcatccaggcctgatcgggctcatcggtcc4860
tccgggtgaacagggtgagaagggcgaccgtggtctccctggcccccagggctcctccgg4920
tcctaagggagaacagggtatcactggtccttctggcccgattgggcctcctgggccccc4980
tggcctgccgggtccgcctggtccaaaaggtgctaagggctcctcgggtccaactggccc5040
gaagggtgaggcaggccacccaggacccccaggccccccgggccccccgggagaggtcat5100
ccagcccctgccaatccaggcatccaggacgcggcggaacatcgacgccagccagctgct5160
ggacgacgggaatggcgagaactacgtggactacgcggacggcatggaagagatcttcgg5220
ctctctcaactctctgaagctggagattgagcagatgaaacggcccctgggcacgcagca5280
gaaccccgcccgcacctgcaaggacctgcagctctgccaccccgacttcccagatggtga5340
atactgggtcgatcctaaccaaggatgctccagggattccttcaaggtttactgcaactt5400
cacagccggggggtcgacatgcgtcttccctgacaagaagtccgaaggggccagaatcac5460
ttcttggcccaaagaaaacccgggctcctggttcagtgaattcaagcgtgggaaactgct5520
ctcctatgtggacgccgagggcaaccctgtgggtgtggtacagatgaccttcctgcggct5580
gctgagcgcctctgcccaccagaacgtcacctaccactgctaccagtcagtggcctggca5640
ggacgcagccacgggcagctacgacaaggccctccgcttcctgggctccaacgacgagga5700
gatgtcctatgacaacaacccctacatccgcgccctggtggacggctgtgctaccaagaa5760
aggctaccagaagacggttctggagatcgacacccccaaagtggagcaggtgcccatcgt5820
ggacatcatgttcaatgacttcggtgaagcgtcacagaaatttggatttgaagtggggcc5880
ggcttgcttcatgggctaggagccgccgagcccgggctcccgagagcaacctcgtgacct5940
cagcatgccattcgttcgtgagtgtcccgtgcacgtcctgaccctggacagtgaaggctt6000
ctccctcccctcccacctgacttcatctacgcctcggcaccacggggtgtgggaccccag6060
cccggagagaacagagggaaggagccgcgcccccacctggagctgaatcacatgacctag6120
ctgcaccccagcgcctgggcccgccccacgctctgtccacacccacgcgccccgggagcg6180
gggccatgcctccagccccccagctcgcccgacccatcctgttcgtgaataggtctcagg6240
ggttgggggagggactgccagatttggacactatatttttttctaaattcaacttgaaga6300
tgtgtatttcccctgaccttcaaaaaatgttccaaggtaagcctcgtaaaggtcatccca6360
ccatcaccaaagcctccgtttttaacaacctccaacacgatccatttagaggccaaatgt6420
cattctgcaggtgccttcccgatggattaaaggtgcttatgtttttgtgagttttaagta6480
aatatttgtattgtattgttataaatgttaagtgtgcctggctttcaatcatgcacggaa6540
acccagtctcagtcccacggacagaatgggcgaggcatggattctgggttgcagtaccgt6600
tctgattagaaataggaagtctccccacccccgccctggccaagaacgtgcaataaattg6660
gaagtttgccccggggcagcaagaatttatgctgccattgaaaagcaggtaccagtgccc6720
cttttcagacagtttttgattcgctctagactttttttttttttaatagggaaaaaattt6780
gataattttcttttttctacatgcacttaagactaaaacacaggtttggattaattttat6840
ttgcttcctttttccgcttttcttcccgcagagcctgatgggagaatgtccagggcaggg6900
aaaccacattttttgtaggtgataactcaatgaaaattggtgcttattttttacacttct6960
ctcttgtggctctcttgtggtgctatctatctgttttaaggtctccttgaaggcgcactg7020
gggaccctggccatgcctcgttctccctgctttctttatcctgttattgcctccacagtc7080
tgttgccaaggactctaagatcaatgcacgtcactttcctttccactgggcaggatagcc7140
aagcacactccctcctgcgctctcccgccccggtgcgtccactcccgagggctgttatga7200
ggactgggttgtgcctacttgatttgaaaacacacacaagcaataaaaagcctcttcctg7260
cattgtctgtggtgtgaccatagcagattatatttggttcctgaatgtttgtggtgctaa7320
tttctgtgtttgttccaagccgttcagtcatgccatgcgctgcctcggtagatggagtaa7380
tgtacaatgaactccatgagtctctccagggctgcctgcagcacgtcttttccaagtagc7440
ctatttggattcccatctcaaatgtcctggatgcgagcgtcagcggctccagagctcggg7500
gcgggtgaggtcccctttggggaaccctttcctggccatcgaggtcggggggctgccgtc7560
tgtgggcaggaggacccgaggggcagccaggaaaggcgatctcttcactgtgaaaagttg7620
cccgggtgcagcgccttttccttctaccatgggaaatgcaggctgggcccttggggtgag7680
cctgcggggctctggtgctgtccccgacccccaccaccaccagaatgcagttccagctta7740
ggaagccacaaacaagccacccaggaggaacaaaacaccgccagcgtggattttccaaat7800
ttccctggaaagtaagtctcgctcttgccaaagaaaagtctggcttggagagtctctgga7860
gcccaggatgccagcatgtgccaatgactgtcaccttcatctcttcaaaagaaaagccat7920
agccgaggactgtcccgcgacccccgtggactgcgtctaggtcatgtgattctgttttca7980
tttctcatcccatccaatttgtccttttctcctgtcattttcttcctctgtggtcccttc8040
aaagttgttataatttgtactgaacttcaaaatgtgtcccgttctccccagaccactcta8100
gccacagtatattgcaataaaattacttcttatatttgcagaaattcttttggtgtaatt8160
ttattttttcctctcaatatatataattggacaaacgctggcaaaaagaaaaaaatggta8220
agcaaaaaacccaagataaagtttcgaggacatcaggccttttgaaatacaatgtcaaat8280
gacacattgtacggtttcaaaaaatccgctagacatgtcataagttttaactgtaatgcc8340
caggaaaggatatcttaaaatattctaaacttgtgtaacaaaggaataattaactgtaat8400
agtttttcaataaatcgagttgggtgtttccaccgtaaa8439
<210>2
<211>1838
<212>prt
<213>智人
<400>2
metaspvalhisthrargtrplysalaargseralaleuargprogly
151015
alaproleuleuproproleuleuleuleuleuleutrpalapropro
202530
proserargalaalaglnproalaaspleuleulysvalleuaspphe
354045
hisasnleuproaspglyilethrlysthrthrglyphecysalathr
505560
argargserserlysglyproaspvalalatyrargvalthrlysasp
65707580
alaglnleuseralaprothrlysglnleutyrproalaseralaphe
859095
progluasppheserileleuthrthrvallysalalyslysglyser
100105110
glnalapheleuvalseriletyrasngluglnglyileglnglnile
115120125
glyleugluleuglyargserprovalpheleutyrgluasphisthr
130135140
glylysproglyprogluasptyrproleupheargglyileasnleu
145150155160
seraspglylystrphisargilealaleuservalhislyslysasn
165170175
valthrleuileleuaspcyslyslyslysthrthrlyspheleuasp
180185190
argserasphisprometileaspileasnglyileilevalphegly
195200205
thrargileleuaspglugluvalphegluglyaspileglnglnleu
210215220
leuphevalserasphisargalaalatyrasptyrcysgluhistyr
225230235240
serproaspcysaspthralavalproaspthrproglnserglnasp
245250255
proasnproaspglutyrtyrthrgluglyaspglygluglygluthr
260265270
tyrtyrtyrglutyrprotyrtyrgluaspprogluaspleuglylys
275280285
gluprothrproserlyslysprovalglualaalalysgluthrthr
290295300
gluvalproglugluleuthrprothrprothrglualaalapromet
305310315320
progluthrsergluglyalaglylysglugluaspvalglyilegly
325330335
asptyrasptyrvalprosergluasptyrtyrthrproserprotyr
340345350
aspaspleuthrtyrglygluglyglugluasnproaspglnprothr
355360365
aspproglyalaglyalagluileprothrserthralaaspthrser
370375380
asnserserasnproalaproproproglygluglyalaaspaspleu
385390395400
gluglygluphethrglugluthrileargasnleuaspgluasntyr
405410415
tyraspprotyrtyraspprothrserserprosergluileglypro
420425430
glymetproalaasnglnaspthriletyrgluglyileglyglypro
435440445
argglyglulysglyglnlysglygluproalaileilegluprogly
450455460
metleuilegluglyproproglyprogluglyproalaglyleupro
465470475480
glyproproglythrmetglyprothrglyglnvalglyaspprogly
485490495
gluargglyproproglyargproglyleuproglyalaaspglyleu
500505510
proglyproproglythrmetleumetleupropheargpheglygly
515520525
glyglyaspalaglyserlysglyprometvalseralaglngluser
530535540
glnalaglnalaileleuglnglnalaargleualaleuargglypro
545550555560
alaglyprometglyleuthrglyargproglyprovalglypropro
565570575
glyserglyglyleulysglygluproglyaspvalglyproglngly
580585590
proargglyvalglnglyproproglyproalaglylysproglyarg
595600605
argglyargalaglyseraspglyalaargglymetproglyglnthr
610615620
glyprolysglyaspargglypheaspglyleualaglyleuprogly
625630635640
glulysglyhisargglyaspproglyproserglyproproglypro
645650655
proglyaspaspglygluargglyaspaspglygluvalglyproarg
660665670
glyleuproglygluproglyproargglyleuleuglyprolysgly
675680685
proproglyproproglyproproglyvalthrglymetaspglygln
690695700
proglyprolysglyasnvalglyproglnglygluproglypropro
705710715720
glyglnglnglyasnproglyalaglnglyleuproglyproglngly
725730735
alaileglyproproglyglulysglyproleuglylysproglyleu
740745750
proglymetproglyalaaspglyproproglyhisproglylysglu
755760765
glyproproglyglulysglyglyglnglyproproglyproglngly
770775780
proileglytyrproglyproargglyvallysglyalaaspglyile
785790795800
argglyleulysglythrlysglyglulysglygluaspglyphepro
805810815
glyphelysglyaspmetglyilelysglyaspargglygluilegly
820825830
proproglyproargglygluaspglyprogluglyprolysglyarg
835840845
glyglyproasnglyaspproglyproleuglyproproglyglulys
850855860
glylysleuglyvalproglyleuproglytyrproglyargglngly
865870875880
prolysglyserileglypheproglypheproglyalaasnglyglu
885890895
lysglyglyargglythrproglylysproglyproargglyglnarg
900905910
glyprothrglyproargglygluargglyproargglyilethrgly
915920925
lysproglyprolysglyasnserglyglyaspglyproalaglypro
930935940
proglygluargglyproasnglyproglnglyprothrglyphepro
945950955960
glyprolysglyproproglyproproglylysaspglyleuprogly
965970975
hisproglyglnargglygluthrglypheglnglylysthrglypro
980985990
proglyproproglyvalvalglyproglnglyprothrglygluthr
99510001005
glyprometglygluargglyhisproglyproproglyproprogly
101010151020
gluglnglyleuproglyleualaglylysgluglythrlysglyasp
1025103010351040
proglyproalaglyleuproglylysaspglyproproglyleuarg
104510501055
glypheproglyaspargglyleuproglyprovalglyalaleugly
106010651070
leulysglyasngluglyproproglyproproglyproalaglyser
107510801085
proglygluargglyproalaglyalaalaglyproileglyilepro
109010951100
glyargproglyproglnglyproproglyproalaglyglulysgly
1105111011151120
alaproglyglulysglyproglnglyproalaglyargaspglyleu
112511301135
glnglyprovalglyleuproglyproalaglyprovalglypropro
114011451150
glygluaspglyasplysglygluileglygluproglyglnlysgly
115511601165
serlysglyasplysglygluglnglyproproglyprothrglypro
117011751180
glnglyproileglyglnproglyproserglyalaaspglyglupro
1185119011951200
glyproargglyglnglnglyleupheglyglnlysglyaspglugly
120512101215
proargglypheproglyproproglyprovalglyleuglnglyleu
122012251230
proglyproproglyglulysglygluthrglyaspvalglyglnmet
123512401245
glyproproglyproproglyproargglyproserglyalaprogly
125012551260
alaaspglyproglnglyproproglyglyileglyasnproglyala
1265127012751280
valglyglulysglygluproglyglualaglygluproglyleupro
128512901295
glygluglyglyproproglyprolysglygluargglyglulysgly
130013051310
gluserglyproserglyalaalaglyproproglyprolysglypro
131513201325
proglyaspaspglyprolysglyserproglyprovalglyphepro
133013351340
glyaspproglyproproglygluproglyproalaglyglnaspgly
1345135013551360
proproglyasplysglyaspaspglygluproglyglnthrglyser
136513701375
proglyprothrglygluproglyproserglyproproglylysarg
138013851390
glyproproglyproalaglyprogluglyargglnglyglulysgly
139514001405
alalysglyglualaglyleugluglyproproglylysthrglypro
141014151420
ileglyproglnglyalaproglylysproglyproaspglyleuarg
1425143014351440
glyileproglyprovalglygluglnglyleuproglyserprogly
144514501455
proaspglyproproglyprometglyproproglyleuproglyleu
146014651470
lysglyaspserglyprolysglyglulysglyhisproglyleuile
147514801485
glyleuileglyproproglygluglnglyglulysglyasparggly
149014951500
leuproglyproglnglyserserglyprolysglygluglnglyile
1505151015151520
thrglyproserglyproileglyproproglyproproglyleupro
152515301535
glyproproglyprolysglyalalysglyserserglyprothrgly
154015451550
prolysglyglualaglyhisproglyproproglyproproglypro
155515601565
proglygluvalileglnproleuproileglnalaserargthrarg
157015751580
argasnileaspalaserglnleuleuaspaspglyasnglygluasn
1585159015951600
tyrvalasptyralaaspglymetglugluilepheglyserleuasn
160516101615
serleulysleugluilegluglnmetlysargproleuglythrgln
162016251630
glnasnproalaargthrcyslysaspleuglnleucyshisproasp
163516401645
pheproaspglyglutyrtrpvalaspproasnglnglycysserarg
165016551660
aspserphelysvaltyrcysasnphethralaglyglyserthrcys
1665167016751680
valpheproasplyslyssergluglyalaargilethrsertrppro
168516901695
lysgluasnproglysertrpphesergluphelysargglylysleu
170017051710
leusertyrvalaspalagluglyasnprovalglyvalvalglnmet
171517201725
thrpheleuargleuleuseralaseralahisglnasnvalthrtyr
173017351740
hiscystyrglnservalalatrpglnaspalaalathrglysertyr
1745175017551760
asplysalaleuargpheleuglyserasnaspgluglumetsertyr
176517701775
aspasnasnprotyrileargalaleuvalaspglycysalathrlys
178017851790
lysglytyrglnlysthrvalleugluileaspthrprolysvalglu
179518001805
glnvalproilevalaspilemetpheasnasppheglyglualaser
181018151820
glnlyspheglyphegluvalglyproalacysphemetgly
182518301835
<210>3
<211>6930
<212>dna
<213>智人
<400>3
gaccgttgcttggcagacactggatggttatgagcctgaacaagctgaaaaggggcagga60
aaagaagtggaggcagcattcttcctatttaaagctgcatcgcttgaaaaaagttttcgc120
agactgtgctggagctggtgctgaaaaagggggtttgcagaggctgccctggggctggtg180
ctgaaagaagagcccacagctgacttcatggtgctacaataacctcagaatctacttttc240
actctcaggagaacccacatgtctaatatttagacatgatggcaaactgggcggaagcaa300
gacctctcctcattcttattgttttattagggcaatttgtctcaataaaagcccaggaag360
aagacgaggatgaaggatatggtgaagaaatagcctgcactcagaatggccagatgtact420
taaacagggacatttggaaacctgccccttgtcagatctgtgtctgtgacaatggagcca480
ttctctgtgacaagatagaatgccaggatgtgctggactgtgccgaccctgtaacgcccc540
ctggggaatgctgtcctgtctgttcacaaacacctggaggtggcaatacaaattttggta600
gaggaagaaagggacaaaagggagaaccaggattagtgcctgttgtaacaggcatacgtg660
gtcgtccaggaccggcaggacctccaggatcacagggaccaagaggagagcgagggccaa720
aaggaagacctggccctcgtggacctcagggaattgatggagaaccaggtgttcctggtc780
aacctggtgctccaggacctcctggacatccgtcccacccaggacccgatggcttgagca840
ggccgttttcagctcaaatggctgggttggatgaaaaatctggacttgggagtcaagtag900
gactaatgcctggctctgtgggtcctgttggcccaaggggaccacagggtttacaaggac960
agcaaggtggtgcaggacctacaggacctcctggtgaacctggtgatcctggaccaatgg1020
gtccgattggttcacgtggaccagagggccctcctggtaaacctggggaagatggtgaac1080
ctggcagaaatggaaatcctggtgaagtgggatttgcaggatctccgggagctcgtggat1140
ttcctggggctcctggtcttccaggtctgaagggtcaccgaggacacaaaggtcttgaag1200
gccctaaaggtgaagttggagcacctggttccaagggtgaagctggccccactggtccaa1260
tgggtgccatgggtcctctgggtccgaggggaatgccaggagagagagggagacttgggc1320
cacagggtgctcctggacaacgaggtgcacatggtatgcctggaaaacctggaccaatgg1380
gtcctcttgggataccaggctcttctggttttccaggaaatcctggaatgaagggagaag1440
caggtcctacaggggcgcgaggccctgaaggtcctcaggggcagagaggtgaaactgggc1500
ccccaggtccagttggctctccaggtcttcctggtgcaataggaactgatggtactcctg1560
gtgccaaaggcccaacgggctctccgggtacctctggtcctcctggctcagcagggcctc1620
ctggatctccaggacctcagggtagcactggtcctcagggaattcgaggccaaccgggtg1680
atccaggagttccaggtttcaaaggagaagctggcccaaaaggggaaccagggccacatg1740
gtattcagggtccgataggcccacccggtgaagaaggcaaaagaggtcccagaggtgacc1800
caggaacagttggtcctccagggccagtgggagaaaggggtgctcctggcaatcgtggtt1860
ttccaggctctgatggtttacctgggccaaagggtgctcaaggagaacggggtcctgtag1920
gttcttcaggacccaaaggaagccagggggatccaggacgtccaggggaacctgggcttc1980
caggtgctcggggtttgacaggaaatcctggtgttcaaggtcctgaaggaaaacttggac2040
ctttgggtgcgccaggggaagatggccgtccaggtcctccaggctccataggaatcagag2100
ggcagcccgggagcatgggccttccaggccccaaaggtagcagtggtgaccctgggaaac2160
ctggagaagcaggaaatgctggagttcctgggcagaggggagctcctggaaaagatggtg2220
aagttggtccttctggtcctgtgggcccgccgggtctagctggtgaaagaggagaacaag2280
gacctccaggccccacaggttttcaggggcttcctggtcctccagggcctcctggagaag2340
gtggaaaaccaggtgatcaaggtgttcctggagatcccggagcagttggcccgttaggac2400
ctagaggagaacgaggaaatcctggggaaagaggagaacctgggataactggactccctg2460
gtgagaagggaatggctggaggacatggtcctgatggcccaaaaggcagtccaggtccat2520
ctgggacccctggagatacaggcccaccaggtcttcaaggtatgccgggagaaagaggaa2580
ttgcaggaactcctggccccaagggtgacagaggtggcataggagaaaaaggtgctgaag2640
gcacagctggaaatgatggtgcaagaggtcttccaggtcctttgggccctccaggtccgg2700
caggtcctactggagaaaagggtgaacctggtcctcgaggtttagttggccctcctggct2760
cccggggcaatcctggttctcgaggtgaaaatgggccaactggagctgttggttttgccg2820
gaccccagggtcctgacggacagcctggagtaaaaggtgaacctggagagccaggacaga2880
agggagatgctggttctcctggaccacaaggtttagcaggatcccctggccctcatggtc2940
ctaatggtgttcctggactaaaaggtggtcgaggaacccaaggtccgcctggtgctacag3000
gatttcctggttctgcgggcagagttggacctccaggccctgctggagctccaggacctg3060
cgggacccctaggggaacccgggaaggagggacctccaggtcttcgtggggaccctggct3120
ctcatgggcgtgtgggagatcgaggaccagctggcccccctggtggcccaggagacaaag3180
gggacccaggagaagatgggcaacctggtccagatggcccccctggtccagctggaacga3240
ccgggcagagaggaattgttggcatgcctgggcaacgtggagagagaggcatgcccggcc3300
taccaggcccagcgggaacaccaggaaaagtaggaccaactggtgcaacaggagataaag3360
gtccacctggacctgtggggcccccaggctccaatggtcctgtaggggaacctggaccag3420
aaggtccagctggcaatgatggtaccccaggacgggatggtgctgttggagaacgtggtg3480
atcgtggagaccctgggcctgcaggtctgccaggctctcagggtgcccctggaactcctg3540
gccctgtgggtgctccaggagatgcaggacaaagaggagatccgggttctcggggtccta3600
taggaccacctggtcgagctgggaaacgtggattacctggaccccaaggacctcgtggtg3660
acaaaggtgatcatggagaccgaggcgacagaggtcagaagggccacagaggctttactg3720
gtcttcagggtcttcctggccctcctggtccaaatggtgaacaaggaagtgctggaatcc3780
ctggaccatttggcccaagaggtcctccaggcccagttggtccttcaggtaaagaaggaa3840
accctgggccacttgggccaattggacctccaggtgtacgaggcagtgtaggagaagcag3900
gacctgagggccctcctggtgagcctggcccacctggccctccgggtccccctggccacc3960
ttacagctgctcttggggatatcatggggcactatgatgaaagcatgccagatccacttc4020
ctgagtttactgaagatcaggcggctcctgatgacaaaaacaaaacggacccaggggttc4080
atgctaccctgaagtcactcagtagtcagattgaaaccatgcgcagccccgatggctcga4140
aaaagcacccagcccgcacgtgtgatgacctaaagctttgccattccgcaaagcagagtg4200
gtgaatactggattgatcctaaccaaggatctgttgaagatgcaatcaaagtttactgca4260
acatggaaacaggagaaacatgtatttcagcaaacccatccagtgtaccacgtaaaacct4320
ggtgggccagtaaatctcctgacaataaacctgtttggtatggtcttgatatgaacagag4380
ggtctcagttcgcttatggagaccaccaatcacctaatacagccattactcagatgactt4440
ttttgcgccttttatcaaaagaagcctcccagaacatcacttacatctgtaaaaacagtg4500
taggatacatggacgatcaagctaagaacctcaaaaaagctgtggttctcaaaggggcaa4560
atgacttagatatcaaagcagagggaaatattagattccggtatatcgttcttcaagaca4620
cttgctctaagcggaatggaaatgtgggcaagactgtctttgaatatagaacacagaatg4680
tggcacgcttgcccatcatagatcttgctcctgtggatgttggcggcacagaccaggaat4740
tcggcgttgaaattgggccagtttgttttgtgtaaagtaagccaagacacatcgacaatg4800
agcaccaccatcaatgaccaccgccattcacaagaactttgactgtttgaagttgatcct4860
gagactcttgaagtaatggctgatcctgcatcagcattgtatatatggtcttaagtgcct4920
ggcctccttatccttcagaatatttattttacttacaatcctcaagttttaattgatttt4980
aaatatttttcaatacaacagtttaggtttaagatgaccaatgacaatgaccacctttgc5040
agaaagtaaactgattgaataaataaatctccgttttcttcaatttatttcagtgtaatg5100
aaaaagttgcttagtatttatgaggaaattcttcttcctggcaggtagcttaaagagtgg5160
ggtatatagagccacaacacatgtttattttgcttggctgcagttgaaaaatagaaatta5220
gtgcccttttgtgacctctcattccaagattgtcaattaaaaatgagtttaaaatgttta5280
acttgtgatcgagacctacatgcatgtcttgatattgtgtaactataatagagactcttt5340
aaggagaatcttaaaaaaaaaaaaacgtttctcactgtcttaaatagaatttttaaatag5400
tatatattcagtggcattttggagaacaaagtgaatttacttcgacttcttaaatttttg5460
taaaagactataagtttagacatctttctcattcaaatttaaagatatctttctcctctt5520
gatcaatctatcaatattgatagaagtcacactagtatataccatttaatacatttacac5580
tttcttatttaagaagatattgaatgcaaaataattgacatatagaactttacaaacata5640
tgtccaaggactctaaattgagactcttccacatgtacaatctcatcatcctgaagccta5700
taatgaagaaaaagatctagaaactgagttgtggagctgactctaatcaaatgtgatgat5760
tggaattagaccatttggcctttgaactttcataggaaaaatgacccaacatttcttagc5820
atgagctacctcatctctagaagctgggatggacttactattcttgtttatattttagat5880
actgaaaggtgctatgcttctgttattattccaagactggagataggcagggctaaaaag5940
gtattattatttttcctttaatgatggtgctaaaattcttcctataaaattccttaaaaa6000
taaagatggtttaatcactaccattgtgaaaacataactgttagacttcccgtttctgaa6060
agaaagagcatcgttccaatgcttgttcactgttcctctgtcatactgtatctggaatgc6120
tttgtaatacttgcatgcttcttagaccagaacatgtaggtccccttgtgtctcaatact6180
ttttttttcttaattgcatttgttggctctattttaatttttttcttttaaaataaacag6240
ctgggaccatcccaaaagacaagccatgcatacaactttggtcatgtatctctgcaaagc6300
atcaaattaaatgcacgcttttgtcatgtcagtggtttttgttttgtgaaattcctttga6360
ccatattagatctatttcatttccaatagtgaaaaggagatgtggtggtatactttgttt6420
gccatttgtttaaaagatacaacggataccttctatcatgtatgtactggcttataaatg6480
aaaatctatctacaacattacccacaaaggcaacatgacaccaattatcactgcctctgc6540
ccttaaaaatgtcagagtagtattattgataaaaagggcaagcaatagatttttcatgac6600
tgaataaactgtaataataaaacatatgtctcaaagtgtatcacatatgaatttagccta6660
attgttttcagtttcattctcaatatttagtttacaacatcattttcccctaaactggtt6720
atattttgacctgtatatcttaaatttgagtatttatatgcctaaatacatgtgtgagtt6780
ttgtttgacttccaagtccaaactataagattatataagttcatatagatgaatcagaaa6840
tatgtggtaatactattaagtcacaaacactaacaatttccaactatagaaataacagtt6900
cttatttggattttgggaatgctaccaata6930
<210>4
<211>1499
<212>prt
<213>智人
<400>4
metmetalaasntrpalaglualaargproleuleuileleuileval
151015
leuleuglyglnphevalserilelysalaglnglugluaspgluasp
202530
gluglytyrglyglugluilealacysthrglnasnglyglnmettyr
354045
leuasnargaspiletrplysproalaprocysglnilecysvalcys
505560
aspasnglyalaileleucysasplysileglucysglnaspvalleu
65707580
aspcysalaaspprovalthrproproglyglucyscysprovalcys
859095
serglnthrproglyglyglyasnthrasnpheglyargglyarglys
100105110
glyglnlysglygluproglyleuvalprovalvalthrglyilearg
115120125
glyargproglyproalaglyproproglyserglnglyproarggly
130135140
gluargglyprolysglyargproglyproargglyproglnglyile
145150155160
aspglygluproglyvalproglyglnproglyalaproglypropro
165170175
glyhisproserhisproglyproaspglyleuserargpropheser
180185190
alaglnmetalaglyleuaspglulysserglyleuglyserglnval
195200205
glyleumetproglyservalglyprovalglyproargglyprogln
210215220
glyleuglnglyglnglnglyglyalaglyprothrglyproprogly
225230235240
gluproglyaspproglyprometglyproileglyserargglypro
245250255
gluglyproproglylysproglygluaspglygluproglyargasn
260265270
glyasnproglygluvalglyphealaglyserproglyalaarggly
275280285
pheproglyalaproglyleuproglyleulysglyhisargglyhis
290295300
lysglyleugluglyprolysglygluvalglyalaproglyserlys
305310315320
glyglualaglyprothrglyprometglyalametglyproleugly
325330335
proargglymetproglygluargglyargleuglyproglnglyala
340345350
proglyglnargglyalahisglymetproglylysproglypromet
355360365
glyproleuglyileproglyserserglypheproglyasnprogly
370375380
metlysglyglualaglyprothrglyalaargglyprogluglypro
385390395400
glnglyglnargglygluthrglyproproglyprovalglyserpro
405410415
glyleuproglyalaileglythraspglythrproglyalalysgly
420425430
prothrglyserproglythrserglyproproglyseralaglypro
435440445
proglyserproglyproglnglyserthrglyproglnglyilearg
450455460
glyglnproglyaspproglyvalproglyphelysglyglualagly
465470475480
prolysglygluproglyprohisglyileglnglyproileglypro
485490495
proglyglugluglylysargglyproargglyaspproglythrval
500505510
glyproproglyprovalglygluargglyalaproglyasnarggly
515520525
pheproglyseraspglyleuproglyprolysglyalaglnglyglu
530535540
argglyprovalglyserserglyprolysglyserglnglyasppro
545550555560
glyargproglygluproglyleuproglyalaargglyleuthrgly
565570575
asnproglyvalglnglyprogluglylysleuglyproleuglyala
580585590
proglygluaspglyargproglyproproglyserileglyilearg
595600605
glyglnproglysermetglyleuproglyprolysglysersergly
610615620
aspproglylysproglyglualaglyasnalaglyvalproglygln
625630635640
argglyalaproglylysaspglygluvalglyproserglyproval
645650655
glyproproglyleualaglygluargglygluglnglyproprogly
660665670
prothrglypheglnglyleuproglyproproglyproproglyglu
675680685
glyglylysproglyaspglnglyvalproglyaspproglyalaval
690695700
glyproleuglyproargglygluargglyasnproglygluarggly
705710715720
gluproglyilethrglyleuproglyglulysglymetalaglygly
725730735
hisglyproaspglyprolysglyserproglyproserglythrpro
740745750
glyaspthrglyproproglyleuglnglymetproglygluarggly
755760765
ilealaglythrproglyprolysglyaspargglyglyileglyglu
770775780
lysglyalagluglythralaglyasnaspglyalaargglyleupro
785790795800
glyproleuglyproproglyproalaglyprothrglyglulysgly
805810815
gluproglyproargglyleuvalglyproproglyserargglyasn
820825830
proglyserargglygluasnglyprothrglyalavalglypheala
835840845
glyproglnglyproaspglyglnproglyvallysglygluprogly
850855860
gluproglyglnlysglyaspalaglyserproglyproglnglyleu
865870875880
alaglyserproglyprohisglyproasnglyvalproglyleulys
885890895
glyglyargglythrglnglyproproglyalathrglypheprogly
900905910
seralaglyargvalglyproproglyproalaglyalaproglypro
915920925
alaglyproleuglygluproglylysgluglyproproglyleuarg
930935940
glyaspproglyserhisglyargvalglyaspargglyproalagly
945950955960
proproglyglyproglyasplysglyaspproglygluaspglygln
965970975
proglyproaspglyproproglyproalaglythrthrglyglnarg
980985990
glyilevalglymetproglyglnargglygluargglymetprogly
99510001005
leuproglyproalaglythrproglylysvalglyprothrglyala
101010151020
thrglyasplysglyproproglyprovalglyproproglyserasn
1025103010351040
glyprovalglygluproglyprogluglyproalaglyasnaspgly
104510501055
thrproglyargaspglyalavalglygluargglyaspargglyasp
106010651070
proglyproalaglyleuproglyserglnglyalaproglythrpro
107510801085
glyprovalglyalaproglyaspalaglyglnargglyaspprogly
109010951100
serargglyproileglyproproglyargalaglylysargglyleu
1105111011151120
proglyproglnglyproargglyasplysglyasphisglyasparg
112511301135
glyaspargglyglnlysglyhisargglyphethrglyleuglngly
114011451150
leuproglyproproglyproasnglygluglnglyseralaglyile
115511601165
proglypropheglyproargglyproproglyprovalglyproser
117011751180
glylysgluglyasnproglyproleuglyproileglyproprogly
1185119011951200
valargglyservalglyglualaglyprogluglyproproglyglu
120512101215
proglyproproglyproproglyproproglyhisleuthralaala
122012251230
leuglyaspilemetglyhistyraspglusermetproaspproleu
123512401245
progluphethrgluaspglnalaalaproaspasplysasnlysthr
125012551260
aspproglyvalhisalathrleulysserleuserserglnileglu
1265127012751280
thrmetargserproaspglyserlyslyshisproalaargthrcys
128512901295
aspaspleulysleucyshisseralalysglnserglyglutyrtrp
130013051310
ileaspproasnglnglyservalgluaspalailelysvaltyrcys
131513201325
asnmetgluthrglygluthrcysileseralaasnproserserval
133013351340
proarglysthrtrptrpalaserlysserproaspasnlysproval
1345135013551360
trptyrglyleuaspmetasnargglyserglnphealatyrglyasp
136513701375
hisglnserproasnthralailethrglnmetthrpheleuargleu
138013851390
leuserlysglualaserglnasnilethrtyrilecyslysasnser
139514001405
valglytyrmetaspaspglnalalysasnleulyslysalavalval
141014151420
leulysglyalaasnaspleuaspilelysalagluglyasnilearg
1425143014351440
pheargtyrilevalleuglnaspthrcysserlysargasnglyasn
144514501455
valglylysthrvalpheglutyrargthrglnasnvalalaargleu
146014651470
proileileaspleualaprovalaspvalglyglythraspglnglu
147514801485
pheglyvalgluileglyprovalcyspheval
14901495
<210>5
<211>6192
<212>dna
<213>智人
<400>5
gcgagtgactgcaccgagcccgagaagtcgccgcgccccgcagccgccccgactggttcc60
ccgccttgcccgtgggccccgccgggatggggaaccgccgggacctgggccagccgcggg120
ccggtctctgcctgctcctggccgcgctgcagcttctgccggggacgcaggccgatcctg180
tggatgtcctgaaggccctgggtgtgcagggaggccaggctggggtccccgaggggcctg240
gcttctgtccccagaggactccagagggtgaccgggcattcagaattggccaggccagca300
cgctcggcatccccacgtgggaactctttccagaaggccactttcctgagaacttctcct360
tgctgatcaccttgcggggacagccagccaatcagtctgtcctgctgtccatttatgatg420
aaaggggtgcccggcagttgggcctggcactggggccagcgctgggtctcctaggtgacc480
ccttccgccccctcccccagcaggtcaacctcacagatggcaggtggcaccgtgtggccg540
tcagcatagatggtgagatggtgaccctggtagctgactgtgaagctcagccccctgttt600
tgggccatggcccccgcttcatcagcatagctggactcactgtgctggggacccaggacc660
ttggggaaaagactttcgagggagacattcaggagctgctgataagcccagatcctcagg720
ctgccttccaggcttgtgagcggtacctccccgactgtgacaacctggcaccggcagcca780
cagtggctccccagggtgaaccagaaacccctcgtcctcggcggaaggggaagggaaaag840
ggaggaagaaagggcgaggtcgcaaggggaagggcaggaaaaagaacaaggaaatttgga900
cctcaagtccacctcctgactccgcagagaaccagacctccactgacatccccaagacag960
agactccagctccaaatctgcctccgacccccacgcctttggtcgtcacctccactgtga1020
ctactggactcaatgccacgatcctagagaggagcttggaccctgacagtggaaccgagc1080
tggggaccctggagaccaaggcagccagggaggatgaagaaggagatgattccaccatgg1140
gccctgacttccgggcagcagaatatccatctcggactcagttccagatctttcctggtg1200
ctggagagaaaggagcaaaaggagagcccgcagtgattgaaaaggggcagcagtttgagg1260
gacctccaggagccccaggaccccaaggggtggttggcccctcaggccctcccggccccc1320
caggattccctggcgaccctggtccaccgggccctgctggcctcccaggaatccccggca1380
ttgatgggatccgaggcccaccgggcactgtgatcatgatgccgttccagtttgcaggcg1440
gctcctttaaaggccccccagtctcattccagcaggcccaggctcaggcagttctgcagc1500
agactcagctctctatgaaaggcccccctggtccagtggggctcactgggcgcccaggcc1560
ctgtgggtctccccgggcatccaggtctgaaaggagaggagggagcagaagggccacagg1620
gtccccgaggcctgcagggacctcatggaccccctggccgagtgggcaagatgggccgcc1680
ctggagcagatggagctcggggcctcccaggggacactggacctaagggtgatcgtggct1740
tcgatggcctccctgggctgcctggtgagaagggccaaaggggtgactttggccatgtgg1800
ggcaacccggtcccccaggagaggatggtgagaggggagcagagggacctccagggccca1860
ctggccaggctggggagccgggtccacgaggactgcttggccccagaggctctcctggcc1920
ccacgggtcgcccgggtgtgactggaattgatggtgctcctggtgccaaaggcaatgtgg1980
gtcctccaggagaaccaggccctccgggacagcagggaaaccatgggtcccagggactcc2040
ccggtccccagggactcattggcactcctggggagaagggtccccctggaaacccaggaa2100
ttccaggcctcccaggatccgatggccctctgggtcacccaggacatgagggccccacgg2160
gagagaaaggggctcagggtccaccagggtcggcaggccctccgggctatcctggacctc2220
ggggagtgaagggcacttcaggcaaccggggcctccagggggagaaaggcgagaagggag2280
aggacggcttcccaggcttcaagggcgatgtggggctcaaaggtgatcaggggaaacccg2340
gagctccaggtccccggggagaggatggtcctgaggggccgaaggggcaggcggggcagg2400
ctggcgaggaggggcccccaggctcagctggggagaagggcaagcttggggtgccaggcc2460
tcccaggttatccaggacgccctggacctaagggatctattggatttcccggtcccctgg2520
gacccataggagagaaagggaagtcgggaaagacagggcagccaggcctggaaggagagc2580
ggggaccaccaggttcccgtggagagagggggcaaccgggtgccacagggcaaccaggcc2640
ccaagggcgatgtgggccaggatggagcccctgggatccctggagaaaagggcctccctg2700
gtctgcaaggccctccaggattccctgggccaaagggcccccctggtcaccaaggtaaag2760
atgggcgaccagggcaccctggacagagaggagaactgggcttccaaggtcagacaggcc2820
cgcctggaccagctggtgtcttaggccctcagggaaagacaggagaagtgggacctctag2880
gtgaaagggggcctccaggcccccctggacctcctggtgaacaaggtcttcctggcctgg2940
aaggcagagagggggccaagggggaactgggaccaccaggaccccttgggaaagaagggc3000
cagctggactcaggggctttcccggccccaaagggggccctggggacccgggacctactg3060
gcttaaagggtgataagggccccccagggcccgtgggggccaatggctcccctggtgagc3120
gcggtcctttgggcccagcaggaggcattggacttcctggccaaagtggcagcgaaggcc3180
ccgttggccctgcaggcaagaaggggtcccggggagaacgtggcccccctggccccactg3240
gcaaagatgggatcccagggcccctggggcctctgggaccccctggagctgctgggcctt3300
ctggcgaggaaggggacaagggggatgtgggtgcccccggacacaaggggagtaaaggcg3360
ataaaggagacgcgggcccacctggacaaccagggatacggggtcctgcaggacacccag3420
gtcccccgggagcagacggggctcaggggcgccggggacccccaggcctctttgggcaga3480
aaggagatgacggagtcagaggctttgtgggggtgattggccctcctggactgcaggggc3540
tgccaggccctccgggagagaaaggggaggtcggagacgtcgggtccatgggtccccatg3600
gagctccaggtcctcggggtccccaaggccccactggatcagagggcactccagggctgc3660
ctggaggagttggtcagccaggcgccgtgggtgagaagggtgagcgaggggacgctggag3720
acccagggcctccaggagccccaggcatcccggggcccaagggagacattggtgaaaagg3780
gggactcaggcccatctggagctgctggacccccaggcaagaaaggtccccctggagagg3840
atggagccaaagggagcgtgggccccacggggctgcccggagatctagggcccccaggag3900
accctggagtttcaggcatagatggttccccaggggagaagggagaccctggtgatgttg3960
ggggaccgggtccgcctggagcttctggggagcccggcgcccccgggccccccggcaaga4020
ggggtccttcaggccacatgggtcgagaaggcagagaaggggagaaaggtgccaaggggg4080
agccaggtcctgatgggcccccagggaggacgggtccaatgggggctagagggccccctg4140
gacgtgtggggcctgagggtcttcgagggatccctggccctgtgggtgaaccaggcctcc4200
tgggagcccctggacagatgggccctcctggccccctggggccctctggcctcccagggc4260
tgaagggagacactggccccaagggggaaaagggccacattggattgatcggtctcattg4320
gccccccgggagaagctggtgagaaaggagatcaggggttgccaggcgtgcagggacccc4380
ctggtcccaagggagaccctggtccccctggtcccattggctctctgggccaccctgggc4440
ccccaggtgtggcgggccctctaggacagaaaggctcaaaagggtctccggggtccatgg4500
gcccccgtggagacactggacctgcaggcccaccaggccccccgggtgcccctgccgagc4560
tgcatgggctgcgcaggcgccggcgcttcgtcccagtcccgcttccagtcgtggagggcg4620
gcctggaggaggtgctggcctcgctcacatcgctgagcttggagctggagcagctgcggc4680
gtcctcccggcactgcggagcgcccgggcctcgtgtgccacgagctgcaccgcaaccacc4740
cgcacctgcctgatggggaatactggattgaccccaaccagggctgcgcgcgggactcgt4800
tcagggttttttgcaacttcacggcgggaggagagacctgcctctatcccgacaagaagt4860
ttgagatcgtgaaattggcctcctggtccaaggaaaagcctggaggctggtatagcacat4920
tccgtcgagggaagaagttctcctacgtggacgccgacgggtccccagtgaatgtcgtgc4980
agctgaacttcctgaaactgctgagtgccacagctcgccagaacttcacctactcctgcc5040
agaatgcagctgcctggctggacgaagccacgggtgactacagccactccgcccgcttcc5100
ttggcaccaatggagaggagctgtctttcaaccagacgacagcagccactgtcagcgtcc5160
cccaggatggctgccggctccggaaaggacagacgaagacccttttcgaattcagctctt5220
ctcgagcgggatttctgcccctgtgggatgtggcggccactgactttggccagacgaacc5280
aaaagtttgggtttgaactgggccccgtctgcttcagcagctgagagtgtccggggtggg5340
agggaccatgagggagccccagaatggggtgcatttggtgctgaggctttgaagccaccg5400
tatttttcgttacctgtgactatggagccaatgggatgtgacttcgctcatcacggacag5460
tcattccttctcctttccagggtgctgggggctggggttccctggcccaagggtccagcc5520
tcctctcaccccattccaggtggcatactgcagtctggctctttctcccctccctcccca5580
cccaagcctcacctccccaccccttgaacccccatgcaatgagcttctaactcagagctg5640
atgaacaaaagcccccccacccccaatgcctgcctcctcactcctccgtcgctgcccttc5700
acaccttttggtgctacccctccccagagttaagcactggatgtctcctgatcccaggct5760
gggacccctacccccaccccctttgatcctttctacttccacggtgaaaggactgaggtc5820
ggactacagagggaagagggacttcccttgactgggttgtgtttcttttcctgcctcagc5880
ccagctctgcaaatcccctccccctgccccccacctccccaggctcaccttgccatgcca5940
ggtggtttggggaccaagatgttgggggggtgaatcaggatcctaatggtgctgccctat6000
ttatacctgggtctgtattaaaagggaaagtcccccctgttgtagatttcatctgcttcc6060
tccttagggaaggctgggatatgatgagagattccagcccaagcctggccccccaccgcc6120
aggccatagggcataatttgcatctcaaatctgagaataaactgatgaactgtgaaaaaa6180
aaaaaaaaaaaa6192
<210>6
<211>1745
<212>prt
<213>智人
<400>6
metglyasnargargaspleuglyglnproargalaglyleucysleu
151015
leuleualaalaleuglnleuleuproglythrglnalaaspproval
202530
aspvalleulysalaleuglyvalglnglyglyglnalaglyvalpro
354045
gluglyproglyphecysproglnargthrprogluglyaspargala
505560
pheargileglyglnalaserthrleuglyileprothrtrpgluleu
65707580
pheprogluglyhispheprogluasnpheserleuleuilethrleu
859095
argglyglnproalaasnglnservalleuleuseriletyraspglu
100105110
argglyalaargglnleuglyleualaleuglyproalaleuglyleu
115120125
leuglyasppropheargproleuproglnglnvalasnleuthrasp
130135140
glyargtrphisargvalalavalserileaspglyglumetvalthr
145150155160
leuvalalaaspcysglualaglnproprovalleuglyhisglypro
165170175
argpheileserilealaglyleuthrvalleuglythrglnaspleu
180185190
glyglulysthrphegluglyaspileglngluleuleuileserpro
195200205
aspproglnalaalapheglnalacysgluargtyrleuproaspcys
210215220
aspasnleualaproalaalathrvalalaproglnglygluproglu
225230235240
thrproargproargarglysglylysglylysglyarglyslysgly
245250255
argglyarglysglylysglyarglyslysasnlysgluiletrpthr
260265270
serserproproproaspseralagluasnglnthrserthraspile
275280285
prolysthrgluthrproalaproasnleuproprothrprothrpro
290295300
leuvalvalthrserthrvalthrthrglyleuasnalathrileleu
305310315320
gluargserleuaspproaspserglythrgluleuglythrleuglu
325330335
thrlysalaalaarggluaspglugluglyaspaspserthrmetgly
340345350
proasppheargalaalaglutyrproserargthrglnpheglnile
355360365
pheproglyalaglyglulysglyalalysglygluproalavalile
370375380
glulysglyglnglnphegluglyproproglyalaproglyprogln
385390395400
glyvalvalglyproserglyproproglyproproglypheprogly
405410415
aspproglyproproglyproalaglyleuproglyileproglyile
420425430
aspglyileargglyproproglythrvalilemetmetprophegln
435440445
phealaglyglyserphelysglyproprovalserpheglnglnala
450455460
glnalaglnalavalleuglnglnthrglnleusermetlysglypro
465470475480
proglyprovalglyleuthrglyargproglyprovalglyleupro
485490495
glyhisproglyleulysglyglugluglyalagluglyproglngly
500505510
proargglyleuglnglyprohisglyproproglyargvalglylys
515520525
metglyargproglyalaaspglyalaargglyleuproglyaspthr
530535540
glyprolysglyaspargglypheaspglyleuproglyleuprogly
545550555560
glulysglyglnargglyasppheglyhisvalglyglnproglypro
565570575
proglygluaspglygluargglyalagluglyproproglyprothr
580585590
glyglnalaglygluproglyproargglyleuleuglyproarggly
595600605
serproglyprothrglyargproglyvalthrglyileaspglyala
610615620
proglyalalysglyasnvalglyproproglygluproglypropro
625630635640
glyglnglnglyasnhisglyserglnglyleuproglyproglngly
645650655
leuileglythrproglyglulysglyproproglyasnproglyile
660665670
proglyleuproglyseraspglyproleuglyhisproglyhisglu
675680685
glyprothrglyglulysglyalaglnglyproproglyseralagly
690695700
proproglytyrproglyproargglyvallysglythrserglyasn
705710715720
argglyleuglnglyglulysglyglulysglygluaspglyphepro
725730735
glyphelysglyaspvalglyleulysglyaspglnglylysprogly
740745750
alaproglyproargglygluaspglyprogluglyprolysglygln
755760765
alaglyglnalaglyglugluglyproproglyseralaglyglulys
770775780
glylysleuglyvalproglyleuproglytyrproglyargprogly
785790795800
prolysglyserileglypheproglyproleuglyproileglyglu
805810815
lysglylysserglylysthrglyglnproglyleugluglygluarg
820825830
glyproproglyserargglygluargglyglnproglyalathrgly
835840845
glnproglyprolysglyaspvalglyglnaspglyalaproglyile
850855860
proglyglulysglyleuproglyleuglnglyproproglyphepro
865870875880
glyprolysglyproproglyhisglnglylysaspglyargprogly
885890895
hisproglyglnargglygluleuglypheglnglyglnthrglypro
900905910
proglyproalaglyvalleuglyproglnglylysthrglygluval
915920925
glyproleuglygluargglyproproglyproproglyproprogly
930935940
gluglnglyleuproglyleugluglyarggluglyalalysglyglu
945950955960
leuglyproproglyproleuglylysgluglyproalaglyleuarg
965970975
glypheproglyprolysglyglyproglyaspproglyprothrgly
980985990
leulysglyasplysglyproproglyprovalglyalaasnglyser
99510001005
proglygluargglyproleuglyproalaglyglyileglyleupro
101010151020
glyglnserglysergluglyprovalglyproalaglylyslysgly
1025103010351040
serargglygluargglyproproglyprothrglylysaspglyile
104510501055
proglyproleuglyproleuglyproproglyalaalaglyproser
106010651070
glyglugluglyasplysglyaspvalglyalaproglyhislysgly
107510801085
serlysglyasplysglyaspalaglyproproglyglnproglyile
109010951100
argglyproalaglyhisproglyproproglyalaaspglyalagln
1105111011151120
glyargargglyproproglyleupheglyglnlysglyaspaspgly
112511301135
valargglyphevalglyvalileglyproproglyleuglnglyleu
114011451150
proglyproproglyglulysglygluvalglyaspvalglysermet
115511601165
glyprohisglyalaproglyproargglyproglnglyprothrgly
117011751180
sergluglythrproglyleuproglyglyvalglyglnproglyala
1185119011951200
valglyglulysglygluargglyaspalaglyaspproglypropro
120512101215
glyalaproglyileproglyprolysglyaspileglyglulysgly
122012251230
aspserglyproserglyalaalaglyproproglylyslysglypro
123512401245
proglygluaspglyalalysglyservalglyprothrglyleupro
125012551260
glyaspleuglyproproglyaspproglyvalserglyileaspgly
1265127012751280
serproglyglulysglyaspproglyaspvalglyglyproglypro
128512901295
proglyalaserglygluproglyalaproglyproproglylysarg
130013051310
glyproserglyhismetglyarggluglyarggluglyglulysgly
131513201325
alalysglygluproglyproaspglyproproglyargthrglypro
133013351340
metglyalaargglyproproglyargvalglyprogluglyleuarg
1345135013551360
glyileproglyprovalglygluproglyleuleuglyalaprogly
136513701375
glnmetglyproproglyproleuglyproserglyleuproglyleu
138013851390
lysglyaspthrglyprolysglyglulysglyhisileglyleuile
139514001405
glyleuileglyproproglyglualaglyglulysglyaspglngly
141014151420
leuproglyvalglnglyproproglyprolysglyaspproglypro
1425143014351440
proglyproileglyserleuglyhisproglyproproglyvalala
144514501455
glyproleuglyglnlysglyserlysglyserproglysermetgly
146014651470
proargglyaspthrglyproalaglyproproglyproproglyala
147514801485
proalagluleuhisglyleuargargargargargphevalproval
149014951500
proleuprovalvalgluglyglyleuglugluvalleualaserleu
1505151015151520
thrserleuserleugluleugluglnleuargargproproglythr
152515301535
alagluargproglyleuvalcyshisgluleuhisargasnhispro
154015451550
hisleuproaspglyglutyrtrpileaspproasnglnglycysala
155515601565
argaspserpheargvalphecysasnphethralaglyglygluthr
157015751580
cysleutyrproasplyslysphegluilevallysleualasertrp
1585159015951600
serlysglulysproglyglytrptyrserthrpheargargglylys
160516101615
lysphesertyrvalaspalaaspglyserprovalasnvalvalgln
162016251630
leuasnpheleulysleuleuseralathralaargglnasnphethr
163516401645
tyrsercysglnasnalaalaalatrpleuaspglualathrglyasp
165016551660
tyrserhisseralaargpheleuglythrasnglyglugluleuser
1665167016751680
pheasnglnthrthralaalathrvalservalproglnaspglycys
168516901695
argleuarglysglyglnthrlysthrleupheglupheserserser
170017051710
argalaglypheleuproleutrpaspvalalaalathraspphegly
171517201725
glnthrasnglnlyspheglyphegluleuglyprovalcyspheser
173017351740
ser
1745
- 還沒有人留言評論。精彩留言會獲得點贊!