一種水稻EPSPS突變體及其編碼基因和應用的制作方法
本發明涉及基因工程技術領域,具體而言,涉及一種水稻EPSPS突變體及其編碼基因和應用。
背景技術:
草甘膦是目前世界上使用的最為廣泛的除草劑之一,至今已使用了近四十年。草甘膦抑制5-烯醇丙酮莽草酸-3-磷酸合成酶(EPSPS)的活性。EPSPS在莽草酸途徑中催化PEP和shikimate-3-phosphate合成EPSP,最終合成芳香族氨基酸色氨酸、苯丙氨酸和酪氨酸。草甘膦導致芳香族胺基酸合成受阻,進而影響植物正常生長,終致死亡。
目前培育抗草甘膦品種的方法是應用基因工程手段將來自細菌的抗草甘膦基因導入農作物中,從而培育出轉基因抗草甘膦作物新品種。1996年開始推廣以來,種植面積迅速增加,至2015年,全球抗草甘膦轉基因作物種植面積已達1.5億公頃,占轉基因作物種植總面積的83%,給農業生產和環境帶來了巨大利益。
但是,目前農業上應用最廣的抗草甘膦基因是來源于農桿菌(Agrobacterium tumefaciens)CP4菌株的CP4EPSPS。雖然先后從微生物中發現了許多能抗草甘膦的EPSPS基因,但這些基因在農作物中尚未得到廣泛應用。這些微生物抗草甘膦基因在農作物中的應用是將這些基因如CP4EPSPS,用轉基因的方法在農作物中表達。由此而產生的轉基因作物雖然有相當大的推廣面積,但由于反轉基因浪潮,轉基因作物在全世界的接受程度任然較低,即使在轉基因作物種植面積最大的美洲,轉基因也主要局限于玉米、大豆、棉花等幾個作物。
技術實現要素:
本發明的目的在于提供一種水稻EPSPS突變體(即5-烯醇丙酮莽草酸-3-磷酸合成酶突變體),其來源水稻,具有草甘膦抗性。
本發明的另一目的在于提供一種編碼基因,其編碼上述的水稻EPSPS突變體。
本發明的另一目的在于提供一種質粒載體,其含有上述的編碼基因。
本發明的另一目的在于提供含有上述質粒載體的重組菌或重組細胞。
本發明的另一目的在于提供上述編碼基因在培育抗草甘膦植物中的應用。
本發明是這樣實現的:
一種水稻EPSPS突變體,其氨基酸序列如SEQ ID NO.1所示。
一種編碼基因,其編碼上述的EPSPS突變體。
一種質粒載體,其含有上述的編碼基因。
含有上述質粒載體的重組菌或重組細胞。
上述的編碼基因在培育抗草甘膦植物中的應用。
本發明提供的一種水稻EPSPS突變體及其編碼基因和應用有益效果是:
本發明提供的水稻EPSPS突變體,其氨基酸序列如SEQ ID NO.1所示,將其與野生型水稻EPSPS的氨基酸序列(SEQ ID NO.3)相比較,具有多個氨基酸殘基突變位點,分別是由N端至C端的第70位氨基酸殘基由A突變為G、第111位氨基酸殘基由G突變為A、第116位氨基酸殘基由P突變為S、第152位氨基酸殘基由Q突變為R、第213位氨基酸殘基由K突變為R、第250位氨基酸殘基由K突變為Q、第252位氨基酸殘基由K突變為E、以及第342位氨基酸殘基由V突變為A,上述8個位點的突變結果使得該水稻EPSPS突變體具有草甘膦抗性,同時保持了自身的生物酶催化活性,轉化該水稻EPSPS突變體的植株或重組菌均能夠正常生長,且該水稻EPSPS突變體來源于水稻,其可應用于培育抗草甘膦植物例如水稻、煙草、大豆、玉米、小麥、棉花和高粱等,具有廣闊的應用前景。
附圖說明
為了更清楚地說明本發明實施例的技術方案,下面將對實施例中所需要使用的附圖作簡單地介紹,應當理解,以下附圖僅示出了本發明的某些實施例,因此不應被看作是對范圍的限定,對于本領域普通技術人員來講,在不付出創造性勞動的前提下,還可以根據這些附圖獲得其他相關的附圖。
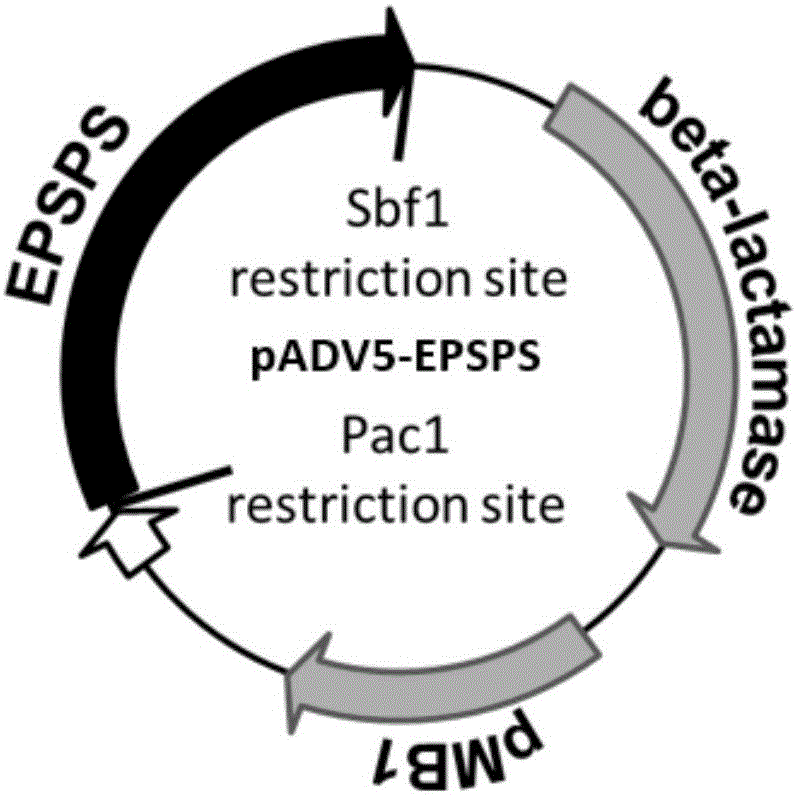
圖1為本發明實施例1提供的pADV5-EPSPS載體的結構示意圖;
圖2為本發明實施例3提供的pBI121-EPSPS載體的結構示意圖;
圖3為本發明實施例1提供的OsEM基因和OsE基因的部分序列對比結果圖;
圖4為本發明實施例4提供的轉OsEM基因和野生型OsE及CP4正對照在不同草甘膦濃度培養基上的生長情況;
圖5為本發明實施例4提供的轉OsEM基因大豆植株的PCR檢測的凝膠電泳結果圖;
圖6為本發明實施例5提供的轉OsEM基因陽性大豆植株經第一次草甘膦處理后的第7天的生長狀況的照片圖;
圖7為本發明實施例5提供的轉OsEM基因陽性大豆植株經第二次草甘膦處理后的第14天的生長狀況的照片圖。
具體實施方式
為使本發明實施例的目的、技術方案和優點更加清楚,下面將對本發明實施例中的技術方案進行清楚、完整地描述。實施例中未注明具體條件者,按照常規條件或制造商建議的條件進行。所用試劑或儀器未注明生產廠商者,均為可以通過市售購買獲得的常規產品。
下面對本發明實施例的水稻EPSPS突變體及其編碼基因和應用進行具體說明。
一方面,本發明提供了一種水稻EPSPS突變體(即5-烯醇丙酮莽草酸-3-磷酸合成酶的突變體),其來源水稻,具有草甘膦抗性。該水稻EPSPS突變體的氨基酸序列如SEQ ID NO.1所示,由454個氨基酸殘基組成。
將其與野生型水稻EPSPS(SEQ ID NO.3)的氨基酸序列相比較,具有多個氨基酸殘基的位點突變。分別是:由N端至C端的第70位氨基酸殘基由A突變為G、第111位氨基酸殘基由G突變為A、第116位氨基酸殘基由P突變為S、第152位氨基酸殘基由Q突變為R、第213位氨基酸殘基由K突變為R、第250位氨基酸殘基由K突變為Q、第252位氨基酸殘基由K突變為E、以及第342位氨基酸殘基由V突變為A(如圖2所示),該多個位點的突變的綜合作用結果使得該水稻EPSPS突變體具有草甘膦抗性,同時保持了自身的生物活性。
再一方面,本發明提供了一種編碼基因,其編碼上述的水稻EPSPS突變體。
根據密碼子的簡并性,在水稻EPSPS突變體的氨基酸序列基礎上能夠很容易地得到該水稻EPSPS突變體的編碼序列。只要能夠編碼出上述水稻EPSPS突變體的編碼序列即屬于本發明的保護范圍。
進一步地,該編碼基因的堿基序列如SEQ ID NO.2所示。
本發明的發明人采用快速定向進化系統,將來源于水稻的野生型EPSPS基因(SEQ ID NO.4)進行錯配PCR,并將產物轉化到敲除EPSPS基因和C-P Lyase基因后的大腸桿菌DH5α,經草甘膦抗性壓篩選后得到了SEQ ID NO.2所示的編碼基因。該快速定向進化系統篩選速度快、效率高。且由于采用的雙敲除的大腸桿菌作為宿主菌,有效地排除了宿主菌內源性基因突變產生的干擾,確保篩選出的突變基因具有相應的草甘膦,有效地排出了假陽性的干擾。
與野生型EPSPS基因(SEQ ID NO.4)相比較,該編碼基因(SEQ ID NO.2)的5’端至3’端的第209位的堿基由C突變為G、第332位的連續兩個堿基由GA突變為CG、第346位由C突變為T、第348位由A突變為C、第455位由A突變為G、第638位由A突變為G、第748位由A突變為C、第755位由A突變為G、第1025位由T突變為C。上述堿基的突變直接導致了水稻EPSPS突變體相應位點的氨基酸殘基突變,賦予了水稻EPSPS突變體的草甘膦抗性。
另一方面,本發明還提供了一種質粒載體,其含有上述的編碼基因。
進一步地,該質粒載體是植物表達載體。
另一方面,本發明還提供了含有上述質粒載體的重組菌或重組細胞。
另一方面,本發明還提供了上述編碼基因在培育抗草甘膦植物中的應用。
進一步地,該應用包括:將所述編碼基因連接至植物表達載體,并轉化目的植物。
其中,轉化目的植物的方法可以根據實際情況選擇,例如農桿菌介導法、基因槍法等。只要是利用本發明提供的編碼基因進行應用培育抗草甘膦植物即屬于本發明的保護范圍。
此外,目的植物的類型可根據實際需要選擇,其可以是水稻、煙草、大豆、玉米、小麥、棉花和高粱中的任意一種。也可以是其他類型的植物或農作物,其均屬于本發明的保護范圍。
此外,容易理解,在本發明提供的上述編碼基因基礎上,本領域的技術人員可以不通過轉基因技術手段來培育抗草甘膦植物,而是以該編碼基因為模板,并修飾目的植物的內源EPSPS基因,例如將內源EPSPS基因的編碼序列修飾為該編碼基因的堿基序列,在目的植物中編碼出與本發明提供的水稻EPSPS突變體一樣的蛋白,使得目的植物具有草甘膦抗性。這樣的應用也屬于本發明的保護范圍。
其中,目的植物包括但不限于水稻、煙草、大豆、玉米、小麥、棉花和高粱。在其他類型的植物中進行上述應用也屬于本發明的保護范圍。
以下結合實施例對本發明的特征和性能作進一步的詳細描述。
實施例1
本實施例提供了的獲取水稻EPSPS基因突變體的方法以及獲得的水稻EPSPS基因突變體,具體步驟如下。
1構建水稻EPSPS基因的突變體庫
1.1采用常規方法將野生型水稻EPSPS基因的mRNA反轉錄成cDNA(如SEQ ID NO.4所示)并克隆到pADV5載體。
1.2用正向引物PV325和反向引物PV323,以連接有水稻EPSPS基因的pADV5載體為模板,進行第一輪錯配PCR。其中,所用引物的堿基序列如下。
PV325:5’-TGAGCGCAACGCAATTAATGTGAG-3’;
PV323:5’-CGTAAGGAGAAAATACCGCATCAGG-3’。
PCR反應體系:25.3μL的H2O、4μL的易錯PCR MIX、4μL的易錯PCR dNTP、4μL的MnCl2、0.8μL的PV325(10μM)、0.8μL的PV323(10μM)、0.1μL的Taq酶、2μL的模板。
PCR反應程序:95℃,30秒;60℃,30秒;72℃,2分鐘;40個循環。PCR產物經1%瓊脂糖電泳,然后切膠回收,得到第一輪PCR產物。
1.3以上述第一輪PCR產物為模板,用正向引物2M1H、反向引物2M1T,進行第二輪PCR。其中,所用引物的堿基序列如下。
2M1H:
5’-ttacgtacgttaattaATGGCGTCCAACGCCGCGGCTGCG-3’
2M1T:
5’-ttacgtacgtcctgcaggTCAGTTCCTGACGAAAGTGCTTAGAACG-3’。
PCR反應體系:31.9μL的H2O、2.5μL的DMSO、5μL的10xPCR buffer、5μL的dNTP、4μL的MgCl2、0.5μL的2M1H(10μM)、0.5μL的2M1T(10μM)、0.1μL的Taq酶、0.5μL的模板(第一輪PCR產物)。
PCR反應程序:95℃,30秒;60℃,30秒;72℃,2分鐘;60個循環
1.4對獲得的PCR產物進行1%瓊脂糖電泳,將與目的條帶大小(1.5kb左右)一致的條帶進行膠回收純化。
1.5對回收的產物進行Pac1和Sbf1雙酶切,然后將其用T4DNA連接酶連接至經同樣雙酶切后的新的pADV5載體上,獲得連接產物。此步驟得到連接產物為攜帶有水稻EPSPS基因突變體的pADV5載體,命名為pADV5-EPSPS,其結構如圖1所示。
1.6轉化雙敲除的大腸桿菌(敲除EPSPS基因和C-P Lyase基因后的大腸桿菌DH5α,命名為EDCE,其制備方法可參考申請號為2016103256926的中國發明專利)
將上述連接產物(pADV5-EPSPS)加入到50μL EDCE感受態細胞中,充分混勻置于冰上30min;42℃熱激90s,冰浴2min后加入LB液體培養基500μL;37℃低速(150r/min)振蕩培養90min。
經過該步驟的處理,攜帶有水稻EPSPS突變基因的pADV5載體轉化到EDCE中,得到轉化產物,也就是水稻EPSPS基因突變體庫。
2抗性篩選
2.1將上述轉化產物低速離心,然后接種于含有不同草甘膦濃度的篩選培養基(5mL)上(篩選培養基相互之間含有的草甘膦濃度有所差別,其含草甘膦的濃度分別是10mM、20mM、50mM,當然草甘膦的濃度可以根據實際情況設定),于37℃、過夜培養。
其中,篩選培養基是以M9為基礎培養基,再添加一定濃度的抗生素Spec(Spectinomycin、奇放線菌素)、Gen(Gentamycin、慶大霉素)、Amp(Ampicillin、氨芐青霉素)以及不同濃度的草甘膦得到的培養基。M9培養基的基礎成分如下:Na2HPO4 13~14g/L、KH2PO4 5.7~6.3g/L、NaCl 0.9~1.1g/L、NH4Cl 1.8~2.2g/L、葡萄糖37~43g/L、MgSO4·7H2O 48~52g/L、CaCl2 21~23g/L。
3測序和序列比對分析
3.1分離在篩選培養基上長出的單克隆抗性菌,然后送樣測序,測pADV5-EPSPS上的EPSPS基因序列,其堿基序列如SEQ ID NO.2所示,即為水稻EPSPS突變基因。將該水稻EPSPS突變基因命名為OsEM基因。其編碼出的5-烯醇丙酮莽草酸-3-磷酸合成酶,命名為水稻EPSPS突變體,氨基酸序列如SEQ ID NO.1所示。
3.2序列比對分析,將得到的OsEM基因與野生型水稻EPSPS基因(SEQ ID NO.4)對比分析。
結果如圖3所示:OsEM基因自5’端至3’端的第209位由C突變為G、第332位的連續兩個堿基由GA突變為CG、第346位由C突變為T、第348位由A突變為C、第455位由A突變為G、第638位由A突變為G、第748位由A突變為C、第755位由A突變為G、第1025位由T突變為C。
其中,第209位由C突變為G,導致水稻EPSPS突變體的第70位氨基酸殘基由A(丙氨酸)突變為G(甘氨酸);
第332位的連續兩個堿基由GA突變為CG,導致水稻EPSPS突變體的第111位氨基酸殘基由G(甘氨酸)突變為A(丙氨酸);
第346位由C突變為T和第348位由A突變為C,導致水稻EPSPS突變體的第116位氨基酸殘基由P(脯氨酸)突變為S(絲氨酸);
第455位由A突變為G,導致水稻EPSPS突變體的第152位氨基酸殘基由Q(谷氨酰胺)突變為R(精氨酸);
第638位由A突變為G,導致水稻EPSPS突變體的第213位氨基酸殘基由K(賴氨酸)突變為R(精氨酸);
第748位由A突變為C,導致水稻EPSPS突變體的第250位氨基酸殘基由K(賴氨酸)突變為Q(谷氨酰胺);
第755位由A突變為G,導致水稻EPSPS突變體的第252位氨基酸殘基由K(賴氨酸)突變為E(谷氨酸);
第1025位由T突變為C,導致水稻EPSPS突變體的第342位氨基酸殘基由V(纈氨酸)突變為A(丙氨酸);共8個氨基酸突變位點(如圖2中的框線所示)。
實施例2
本實施例以轉化了野生型水稻EPSPS基因(命名為OsE基因,SEQ ID NO.3)的EDCE作為對照組,以實施例1中的步驟3.1得到的含OsEM基因(SEQ ID NO.2)的單克隆抗性菌為實驗組,檢測OsEM基因在細菌中的抗草甘膦能力。
將對照組和實驗組分別接種至含0mM、5mM、10mM、20mM、50mM、75mM、100mM濃度草甘膦的培養基(篩選培養基)中,觀察各組的菌落生長狀況,用生長飽和指數來表示(飽和指數=0,沒有生長;飽和指數=1,少量生長;飽和指數=2,生長至半飽和;飽和指數=3,旺盛生長,但還有生長余地;飽和指數=4,快速生長,細菌在培養基中已達最高(飽和)濃度或者說生長已經達到極限)。結果如表1所示。
表1.轉化OsEM基因的實驗組和轉化OsE基因的對照組在含不同濃度草甘膦培養基中的生長飽和指數
表1的結果顯示,在含0mM草甘膦的培養基上實驗組(含OsEM基因)和對照組(含OsE基因)均能正常生長(飽和指數均為4);但在含5mM、10mM、20mM、50mM、75mM、100mM草甘膦的培養基上,對照組不能生長(飽和指數為0),而實驗組能正常生長(飽和指數為4)。由此表明,實施例1得到的OsEM基因(SEQ ID NO.2)能夠賦予EPSPS和C-P Lyase缺陷型的大腸桿菌草甘膦抗性,使其在高達100mM草甘膦的培養基中保持生長狀態。
此外,本實例還用了目前農業上轉基因作物中應用最廣的抗草甘膦CP4-EPSPS基因作為正對照,將分別轉化有OsE基因(負對照組),OsEM基因(實驗組)和CP4-EPSPS基因(正對照組)的大腸桿菌分別接種在含0mM、1mM、50mM、100mM濃度草甘膦的固體培養基上,觀察各組的菌落生長狀況。結果如圖3所示。
圖3的結果顯示,在含0mM草甘膦的培養基上實驗組(含OsEM基因)和負對照組(含OsE基因)及正對照組(含CP4基因)均能正常生長(有明顯白色菌斑);但在含1mM、50mM草甘膦的培養基上,負對照組(OsE)不能生長,而實驗組(OsEM)和正對照(CP4)能正常生長;在含100mM草甘膦的培養基上,負對照組和正對照均不能生長,而實驗組(OsEM)能正常生長。由此表明,實施例1得到水稻EPSPS突變基因(OsEM基因、SEQ ID NO.2)能夠賦予EPSPS和C-P Lyase缺陷型的大腸桿菌草甘膦抗性,使其在高達100mM草甘膦的培養基中保持生長狀態,其草甘膦抗性超過CP4-EPSPS基因。
實施例3
本實施例提供了上述OsEM基因在培育抗草甘膦煙草中的應用。其包括如下步驟。
1制備農桿菌菌液
1.1按常規方法提取由實施例1得到的單克隆抗性菌的質粒(含OsEM基因)。將其用Pac1和Sbf1雙酶切,回收小片段,然后用T4DNA連接酶將小片段連接至經同樣雙酶切后的pBI121載體上,得到pBI121-EPSPS載體,其結構如圖2所示。
1.2將pBI121-EPSPS載體轉化GV3101(根癌農桿菌、Agrobactrium tumefaciens)感受態細胞,挑取單克隆進行菌落PCR檢測,得到陽性菌株;再將陽性菌株接種到1mL的含50μg·mL-1卡那霉素和50μg·mL-1利福平的YEP培養中擴繁,保存于-80℃,或用于后續實驗。
2煙草轉化
2.1吸取上述保存的農桿菌液10μL,接種到1ml的含50μg·mL-1利福平和50μg·mL-1卡那霉素的YEP液體培養基中,28℃,200r/min培養至OD600為0.6-0.8,再將該菌液擴大培養至OD600約為0.5。用MS液體培養液(MS1培養基不加瓊脂)稀釋該菌液并加入20mg/L的AS,作為浸染液。
2.2將煙草種子(野生型“Wisconsin38”)播種在無菌培養基中,選取25-30天的煙草小苗葉片作為轉化外植體。
2.3將葉片除去葉脈和葉邊緣,切割成0.4×0.4cm2方塊,于浸染液中浸泡5-10min,期間不斷搖動浸染液,使菌體與葉片充分接觸。
2.6用鑷子將葉片取出置于無菌濾紙上,待菌液被吸干后將葉片轉至MS1共培養基中,28℃暗培養48h。
2.7將葉片轉至MS2誘導分化培養基中進行選擇培養。15-20天轉接一次,3-4周后有綠芽長出。待芽長到1-2cm高時,切下不定芽,轉入MS3生根篩選培養基進行生根。
待誘導出強壯的根后,取出幼苗,洗凈根部培養基,移栽至育苗土中,幼苗成活,得到轉OsEM基因煙草植株,然后移栽至大田或者溫室或用于后續分析。
3轉基因植株的檢測
采用PCR法檢測出轉OsEM基因煙草植株,根據OsEM基因的序列和煙草內參基因(NtA)設計正反向檢測引物,序列如下:
針對OsEM基因:
OSES53:5’-GCAAGTTGTCCACCACTGTTGTGCC-3’;
ATC5:5’-CTGCAATGGTGTGCAGAACCCATC-3’;擴增片段大小為300bp;
針對NtA內參基因:
NtAct5:5’-CAGGTATTGTGTTGGATTCCGGCG-3’;
NtAct3:5’-CACTGTACTTGCGTTCAGGAGGCG-3’,擴增片段大小為571bp。
分別提取本實施例中的步驟2.7得到轉OsEM基因煙草植株的基因組DNA,均一化至100ng/μL,作為。
PCR檢測體系:10μL 2×TsINGKe,2μL引物混合物(10μmol/L的OSES53、ATC5、NtAct5、NtAct3,各0.5μL),1μL基因組DNA模板(100ng/μL),7μL ddH2O。
PCR檢測程序:94℃,3min;94℃,30s;62℃,30s;72℃,45s;30個循環;72℃,10min;12℃保持。
PCR擴增產物1.5%瓊脂糖凝膠電泳,在300bp位置和571bp位置有條帶的則為轉OsEM基因陽性煙草植株。
實施例4
本實施例提供了上述OsEM基因在培育抗草甘膦大豆中的應用。包括如下步驟。
1OsEM基因轉化大豆
1.1種子消毒和萌發:大豆種子(野生型William 82)采用氯氣干法滅菌,選出干凈飽滿的種子,100粒/皿,放入干燥器,100ml漂白液加8ml鹽酸(12N),消毒時間20小時。使用時取出滅菌后的大豆種子,放入萌發培養基(蔗糖20g/L+GELLAN GUM POWDER),在25℃,暗中培養2天。
1.2侵染與共培養:培養由實施例3步驟1得到的農桿菌菌液至OD600為0.5-1.0時,將其倒入50ml離心管,離心(5000rpm、10min)。去上清,加入浸染液(B5大量、微量及有機+蔗糖30g/L+MES 3.9g/L+GA3 0.25mg/L+6-BA 1.67mg/L+L-Cys 400mg/L+AS 30mg/L)進行重懸,保持0D600在0.5-1.0,得到用于后續浸染的農桿菌重懸液。將吸脹的大豆種子從萌發培養基轉移到無菌培養皿中,沿著種臍用手術刀縱向切割種子,將子葉和下胚軸均勻分開兩瓣,去除種皮后,作為外植體備用。
1.3將農桿菌重懸液倒入潔凈的無菌培養皿中,放入大約200個外植體,室溫侵染1小時。將外植體取,盾片朝下,置于5ml共培養基(B5大量、微量及有機+蔗糖30g/L+MES 3.9g/L+GA30.25mg/L+6-BA 1.67mg/L+L-Cys 400mg/L+DTT 154.2mg/L+AS 30mg/L)上(共培養基上接種的外植體數量可根據情況調整),在25℃條件下,共培養5天。
1.4篩選與再生:將共培養后的外植體轉接到芽誘導培養基(B5大量、微量及有機+蔗糖30g/L+MES 0.59g/L+GELLAN GUM POWDER+6-BA 1.67mg/L+cefotaxime 400mg/L+Glyphosate 15mg/L),于25℃,光周期16/8h、培養3周。
1.5新生芽長出后,轉接至芽伸長培養基(B5大量、微量及有機+蔗糖30g/L+MES 0.59g/L+GELLAN GUM POWDER+6-BA 1.67mg/L+cefotaxime 400mg/L+Glyphosate 15mg/L),于25℃,光周期16/8h、光照培養3-5周。
1.6將伸長3-4cm的幼芽切下,轉接到生根培養基(1/2MS大量、微量、鐵鹽、維他命+蔗糖20g/L+MES 0.59g/L+GELLAN GUM POWDER+IAA 0.2/L),于25℃,光周期16/8h、光照培養至生根;待誘導出強壯的根后,取出幼苗,洗凈根部培養基,移栽至育苗土中,幼苗成活,得到轉OsEM基因大豆植株,然后移栽至大田或者溫室或用于后續分析。
2轉基因植株的檢測
采用PCR法檢測出轉OsEM基因大豆植株,OsEM基因的檢測引物同實施例3(OSES53和ATC5),大豆內參基因(GmC)的正反向檢測引物,序列如下:
針對GmC內參基因:
GmCons6F:5’-CATGCTTTGGCTGTTGGAGATTTG-3’;
GmCons6R:5’-GAACCTGATGGTGTTGGTAGAGGCG-3’;片段大小為658bp。
分別提取上述步驟1.6得到轉OsEM基因大豆植株的基因組DNA(分別編號),均一化至100ng/μL后作為模板。
PCR檢測體系:10μL 2×TsINGKe,2μL引物混合物(10μmol/L的OSES53、ATC5、GmCons6F、GmCons6R,各2μL),1μL基因組DNA(100ng/μL),7μL ddH2O。
PCR檢測程序:94℃,3min;94℃,30s;62℃,30s;72℃,45s;30個循環;72℃,10min;12℃保持。
PCR擴增產物1.5%瓊脂糖凝膠電泳,結果如圖4所示,在300bp位置和658bp位置有條帶的則為轉OsEM基因陽性大豆植株。
圖4的結果顯示(圖中:M為Maeker、Pos為陽性對照、WT為野生型植株、192、193、194、195、197、198、199以及203為不同轉OsEM基因大豆植株編號),編號為192、193、194、195、197、198、199以及203的轉OsEM基因大豆植株有兩條帶,呈陽性,其為轉OsEM基因陽性大豆植株。
實施例5
本實施例以野生型大豆植株為對照,以實施例4得到的轉OsEM基因陽性大豆植株作為實驗組,驗證OsEM基因在植株中的草甘膦抗性。實驗方法如下。
1將生長狀況一致,株高10-20cm的轉OsEM基因陽性大豆植株與3株大小相似的野生型大豆植株編號后隨機混合,均勻地排放在同一實驗區域(避免葉片重疊)。
2計算實驗組和對照組的占地區域面積,根據區域面積,按1060克/公頃(0.106g/m2)劑量來噴施草甘膦。
3用市售的41%草甘膦銨鹽農達,根據上述的噴灑濃度(0.106g/m2),取相應體積的草甘膦銨鹽農達,然后用20倍體積水稀釋后,均勻噴施在實驗組和對照組的植株上。
4待葉面干燥后,將實驗組和對照組的植株移入溫室培養。
5拍照、記錄植株的生長狀況。
生長狀況的統計標準為:如果植株沒有任何的草甘膦損傷現象,生長正常,為高草甘膦抗性植株,用“+++”表示;如果植株的葉片顏色有些發黃,生長略為緩慢,為中草甘膦抗性植株,用“++”表示;如果植株的部分葉片枯萎,生長非常緩慢,為低草甘膦抗性植株,用“+”表示;如果植株枯萎、死亡,為非草甘膦抗性植株(不具有草甘膦抗性),用“-”表示。
噴灑草甘膦后,于第7天觀察并拍照記錄各組植株的生長狀況如圖6所示,并統計出各種抗性的植株數量,結果見表2。
然后將生長良好的植株移栽至大盆中生長,于第21天進行第二次噴灑草甘膦(濃度為1.06g/m2)處理,2周后再拍照,記錄各組植株的生長狀況,結果如7所示。
表2.本實施例的實驗組和對照組經第一次草甘膦處理后第7天的生長狀況統計結果
注:表中的數字表示植株數量
圖6(圖中:1、2、3為野生型植株,a、b、c為轉基因植株)和表2的結果顯示,對照組中,3株野生型大豆植株均表現出不具有草甘膦抗性,而實驗組中,大部分的轉OsE基因陽性大豆植株都具有草甘膦抗性,其中,有4株表現出高草甘膦抗性(如圖6中的a、b、c所示),1株表現出中草甘膦抗性,3株表現出低草甘膦抗性。
由圖7的結果顯示(圖中:a、b和c為轉OsE基因陽性大豆植株、d為野生型大豆植株),實驗組的轉OsE基因陽性大豆植株經1.06g/m2草甘膦處理后能夠持續生長、表現出生長茂盛、葉片持綠度高、植株高等生長旺盛的態勢(如圖7中a和b所示);抗性不強的轉OsE基因陽性大豆植株(如圖7中c所示)表現出生長緩慢和葉片發黃;對照組的野生型大豆植株則枯萎、死亡(如圖7中的d所示)。
以上結果表明,該OsE基因能夠不僅賦予大腸桿菌抗100mM草甘膦濃度的抗性,還能夠賦予大豆植株抗1.06g/m2草甘膦濃度的抗性,該OsE基因的抗性遠遠地高于現有的抗草甘膦基因,其表現出優異草甘膦抗性,同時轉化該OsE基因的大腸桿菌和大豆植株均能正常生長,說明OsE基因編碼的水稻EPSPS突變體的生物酶活性正常。
綜上,相較于野生型水稻EPSPS(SEQ ID NO.3)及其編碼基因(SEQ ID NO.4),本發明提供的水稻EPSPS突變體(SEQ ID NO.1)及其編碼基因(SEQ ID NO.2)具有更高的草甘膦抗性和完整的生物酶活性;此外,該編碼基因(SEQ ID NO.2)來源于水稻品種本身而不是微生物,其可適于轉化各種植物品種例如水稻、煙草、大豆、玉米、棉花、高粱、小麥等植物,其具有更廣闊的適用范圍。同時,根據本發明提供的水稻EPSPS突變基因的序列,可以將其用于培育新的抗草甘膦的水稻品種中(非轉基因方法),獲得抗草甘膦的非轉基因常規品種,提高新品種的公眾接受程度。
以上所述僅為本發明的優選實施例而已,并不用于限制本發明,對于本領域的技術人員來說,本發明可以有各種更改和變化。凡在本發明的精神和原則之內,所作的任何修改、等同替換、改進等,均應包含在本發明的保護范圍之內。
SEQUENCE LISTING
<110> 四川天豫興禾生物科技有限公司
<120> 一種水稻EPSPS突變體及其編碼基因和應用
<160> 4
<170> PatentIn version 3.5
<210> 1
<211> 454
<212> PRT
<213> 人工序列
<400> 1
Met Ala Ser Asn Ala Ala Ala Ala Ala Ala Lys Ala Glu Glu Ile Val
1 5 10 15
Leu Gln Pro Ile Arg Glu Ile Ser Gly Ala Val Gln Leu Pro Gly Ser
20 25 30
Lys Ser Leu Ser Asn Arg Ile Leu Leu Leu Ser Ala Leu Ser Glu Gly
35 40 45
Thr Thr Val Val Asp Asn Leu Leu Asn Ser Glu Asp Val His Tyr Met
50 55 60
Leu Glu Ala Leu Lys Gly Leu Gly Leu Ser Val Glu Ala Asp Lys Val
65 70 75 80
Ala Lys Arg Ala Val Val Val Gly Cys Gly Gly Lys Phe Pro Val Glu
85 90 95
Lys Asp Ala Lys Glu Glu Val Gln Leu Phe Leu Gly Asn Ala Ala Thr
100 105 110
Ala Met Arg Ser Leu Thr Ala Ala Val Thr Ala Ala Gly Gly Asn Ala
115 120 125
Thr Tyr Val Leu Asp Gly Val Pro Arg Met Arg Glu Arg Pro Ile Gly
130 135 140
Asp Leu Val Val Gly Leu Lys Arg Leu Gly Ala Asp Val Asp Cys Phe
145 150 155 160
Leu Gly Thr Glu Cys Pro Pro Val Arg Val Lys Gly Ile Gly Gly Leu
165 170 175
Pro Gly Gly Lys Val Lys Leu Ser Gly Ser Ile Ser Ser Gln Tyr Leu
180 185 190
Ser Ala Leu Leu Met Ala Ala Pro Leu Ala Leu Gly Asp Val Glu Ile
195 200 205
Glu Ile Ile Asp Arg Leu Ile Ser Ile Pro Tyr Val Glu Met Thr Leu
210 215 220
Arg Leu Met Glu Arg Phe Gly Val Lys Ala Glu His Ser Asp Ser Trp
225 230 235 240
Asp Arg Phe Tyr Ile Lys Gly Gly Gln Gln Tyr Glu Ser Pro Gly Asn
245 250 255
Ala Tyr Val Glu Gly Asp Ala Ser Ser Ala Ser Tyr Phe Leu Ala Gly
260 265 270
Ala Ala Ile Thr Gly Gly Thr Val Thr Val Gln Gly Cys Gly Thr Thr
275 280 285
Ser Leu Gln Gly Asp Val Lys Phe Ala Glu Val Leu Glu Met Met Gly
290 295 300
Ala Lys Val Thr Trp Thr Asp Thr Ser Val Thr Val Thr Gly Pro Pro
305 310 315 320
Arg Glu Pro Tyr Gly Lys Lys His Leu Lys Ala Val Asp Val Asn Met
325 330 335
Asn Lys Met Pro Asp Ala Ala Met Thr Leu Ala Val Val Ala Leu Phe
340 345 350
Ala Asp Gly Pro Thr Ala Ile Arg Asp Val Ala Ser Trp Arg Val Lys
355 360 365
Glu Thr Glu Arg Met Val Ala Ile Arg Thr Glu Leu Thr Lys Leu Gly
370 375 380
Ala Ser Val Glu Glu Gly Pro Asp Tyr Cys Ile Ile Thr Pro Pro Glu
385 390 395 400
Lys Leu Asn Ile Thr Ala Ile Asp Thr Tyr Asp Asp His Arg Met Ala
405 410 415
Met Ala Phe Ser Leu Ala Ala Cys Ala Asp Val Pro Val Thr Ile Arg
420 425 430
Asp Pro Gly Cys Thr Arg Lys Thr Phe Pro Asn Tyr Phe Asp Val Leu
435 440 445
Ser Thr Phe Val Arg Asn
450
<210> 2
<211> 1365
<212> DNA
<213> 人工序列
<400> 2
atggcgtcca acgccgcggc tgcggcggcg aaggcggagg agatcgtgct ccagcccatc 60
agggagatct ccggggcggt tcagctgcca gggtccaagt cgctctccaa caggatcctc 120
ctcctctccg ccctctccga gggcacaaca gtggtggaca acttgctgaa cagtgaggat 180
gttcactaca tgcttgaggc cctgaaaggc ctcgggctct ctgtggaagc agataaagtt 240
gcaaaaagag ctgtagtcgt tggctgtggt ggcaagtttc ctgttgagaa ggatgcgaaa 300
gaggaagtgc aactcttctt ggggaacgct gcgactgcaa tgcgatcctt gacagcagcc 360
gtgactgctg ctggtggaaa tgcaacttat gtgcttgatg gagtgccacg aatgagggag 420
agaccgattg gtgacttggt tgtcgggttg aaacgacttg gtgcggatgt cgactgtttc 480
cttggcactg aatgcccacc tgttcgtgtc aagggaattg gaggacttcc tggtggcaag 540
gttaagctct ctggttccat cagcagtcag tacttgagtg ccttgctgat ggctgctcct 600
ttggcccttg gggatgtgga gatcgaaatc attgacagac taatctccat tccttacgtt 660
gaaatgacat tgagattgat ggagcgtttt ggtgtgaagg cagagcattc tgatagttgg 720
gacagattct atattaaggg agggcagcag tacgaatctc ctggaaatgc ctatgttgaa 780
ggtgatgcct caagcgcgag ctatttcttg gctggtgctg caatcactgg aggcactgtg 840
acagttcaag gttgtggtac gaccagtttg cagggtgatg tcaaatttgc tgaggtactt 900
gagatgatgg gagcaaaggt tacatggact gacaccagtg taaccgtaac tggtccacca 960
cgtgagcctt atgggaagaa acacctgaaa gctgttgatg tcaacatgaa caaaatgcct 1020
gatgctgcca tgacccttgc cgttgttgca ctcttcgctg atggtccaac tgctatcaga 1080
gatgtggctt cctggagagt aaaggaaacc gaaaggatgg ttgcaattcg gaccgagcta 1140
acaaagctgg gagcatcggt tgaagaaggt cctgactact gcatcatcac cccaccggag 1200
aagctgaaca tcacggcaat cgacacctac gatgatcaca ggatggccat ggccttctcc 1260
ctcgctgcct gcgccgacgt gcccgtgacg atcagggacc ctggttgcac ccgcaagacc 1320
ttccccaact acttcgacgt tctaagcact ttcgtcagga actga 1365
<210> 3
<211> 454
<212> PRT
<213> 水稻(Oryza sativa)
<400> 3
Met Ala Ser Asn Ala Ala Ala Ala Ala Ala Lys Ala Glu Glu Ile Val
1 5 10 15
Leu Gln Pro Ile Arg Glu Ile Ser Gly Ala Val Gln Leu Pro Gly Ser
20 25 30
Lys Ser Leu Ser Asn Arg Ile Leu Leu Leu Ser Ala Leu Ser Glu Gly
35 40 45
Thr Thr Val Val Asp Asn Leu Leu Asn Ser Glu Asp Val His Tyr Met
50 55 60
Leu Glu Ala Leu Lys Ala Leu Gly Leu Ser Val Glu Ala Asp Lys Val
65 70 75 80
Ala Lys Arg Ala Val Val Val Gly Cys Gly Gly Lys Phe Pro Val Glu
85 90 95
Lys Asp Ala Lys Glu Glu Val Gln Leu Phe Leu Gly Asn Ala Gly Thr
100 105 110
Ala Met Arg Pro Leu Thr Ala Ala Val Thr Ala Ala Gly Gly Asn Ala
115 120 125
Thr Tyr Val Leu Asp Gly Val Pro Arg Met Arg Glu Arg Pro Ile Gly
130 135 140
Asp Leu Val Val Gly Leu Lys Gln Leu Gly Ala Asp Val Asp Cys Phe
145 150 155 160
Leu Gly Thr Glu Cys Pro Pro Val Arg Val Lys Gly Ile Gly Gly Leu
165 170 175
Pro Gly Gly Lys Val Lys Leu Ser Gly Ser Ile Ser Ser Gln Tyr Leu
180 185 190
Ser Ala Leu Leu Met Ala Ala Pro Leu Ala Leu Gly Asp Val Glu Ile
195 200 205
Glu Ile Ile Asp Lys Leu Ile Ser Ile Pro Tyr Val Glu Met Thr Leu
210 215 220
Arg Leu Met Glu Arg Phe Gly Val Lys Ala Glu His Ser Asp Ser Trp
225 230 235 240
Asp Arg Phe Tyr Ile Lys Gly Gly Gln Lys Tyr Lys Ser Pro Gly Asn
245 250 255
Ala Tyr Val Glu Gly Asp Ala Ser Ser Ala Ser Tyr Phe Leu Ala Gly
260 265 270
Ala Ala Ile Thr Gly Gly Thr Val Thr Val Gln Gly Cys Gly Thr Thr
275 280 285
Ser Leu Gln Gly Asp Val Lys Phe Ala Glu Val Leu Glu Met Met Gly
290 295 300
Ala Lys Val Thr Trp Thr Asp Thr Ser Val Thr Val Thr Gly Pro Pro
305 310 315 320
Arg Glu Pro Tyr Gly Lys Lys His Leu Lys Ala Val Asp Val Asn Met
325 330 335
Asn Lys Met Pro Asp Val Ala Met Thr Leu Ala Val Val Ala Leu Phe
340 345 350
Ala Asp Gly Pro Thr Ala Ile Arg Asp Val Ala Ser Trp Arg Val Lys
355 360 365
Glu Thr Glu Arg Met Val Ala Ile Arg Thr Glu Leu Thr Lys Leu Gly
370 375 380
Ala Ser Val Glu Glu Gly Pro Asp Tyr Cys Ile Ile Thr Pro Pro Glu
385 390 395 400
Lys Leu Asn Ile Thr Ala Ile Asp Thr Tyr Asp Asp His Arg Met Ala
405 410 415
Met Ala Phe Ser Leu Ala Ala Cys Ala Asp Val Pro Val Thr Ile Arg
420 425 430
Asp Pro Gly Cys Thr Arg Lys Thr Phe Pro Asn Tyr Phe Asp Val Leu
435 440 445
Ser Thr Phe Val Arg Asn
450
<210> 4
<211> 1365
<212> DNA
<213> 水稻(Oryza sativa)
<400> 4
atggcgtcca acgccgcggc tgcggcggcg aaggcggagg agatcgtgct ccagcccatc 60
agggagatct ccggggcggt tcagctgcca gggtccaagt cgctctccaa caggatcctc 120
ctcctctccg ccctctccga gggcacaaca gtggtggaca acttgctgaa cagtgaggat 180
gttcactaca tgcttgaggc cctgaaagcc ctcgggctct ctgtggaagc agataaagtt 240
gcaaaaagag ctgtagtcgt tggctgtggt ggcaagtttc ctgttgagaa ggatgcgaaa 300
gaggaagtgc aactcttctt ggggaacgct ggaactgcaa tgcgaccatt gacagcagcc 360
gtgactgctg ctggtggaaa tgcaacttat gtgcttgatg gagtgccacg aatgagggag 420
agaccgattg gtgacttggt tgtcgggttg aaacaacttg gtgcggatgt cgactgtttc 480
cttggcactg aatgcccacc tgttcgtgtc aagggaattg gaggacttcc tggtggcaag 540
gttaagctct ctggttccat cagcagtcag tacttgagtg ccttgctgat ggctgctcct 600
ttggcccttg gggatgtgga gatcgaaatc attgacaaac taatctccat tccttacgtt 660
gaaatgacat tgagattgat ggagcgtttt ggtgtgaagg cagagcattc tgatagttgg 720
gacagattct atattaaggg agggcagaag tacaaatctc ctggaaatgc ctatgttgaa 780
ggtgatgcct caagcgcgag ctatttcttg gctggtgctg caatcactgg aggcactgtg 840
acagttcaag gttgtggtac gaccagtttg cagggtgatg tcaaatttgc tgaggtactt 900
gagatgatgg gagcaaaggt tacatggact gacaccagtg taaccgtaac tggtccacca 960
cgtgagcctt atgggaagaa acacctgaaa gctgttgatg tcaacatgaa caaaatgcct 1020
gatgttgcca tgacccttgc cgttgttgca ctcttcgctg atggtccaac tgctatcaga 1080
gatgtggctt cctggagagt aaaggaaacc gaaaggatgg ttgcaattcg gaccgagcta 1140
acaaagctgg gagcatcggt tgaagaaggt cctgactact gcatcatcac cccaccggag 1200
aagctgaaca tcacggcaat cgacacctac gatgatcaca ggatggccat ggccttctcc 1260
ctcgctgcct gcgccgacgt gcccgtgacg atcagggacc ctggttgcac ccgcaagacc 1320
ttccccaact acttcgacgt tctaagcact ttcgtcagga actga 1365
- 還沒有人留言評論。精彩留言會獲得點贊!