一種調控植物體內生長素平衡的EHD1蛋白及其應用的制作方法
本發明屬于生物
技術領域:
,具體涉及一種調控植物體內生長素平衡的EHD1蛋白及其應用。
背景技術:
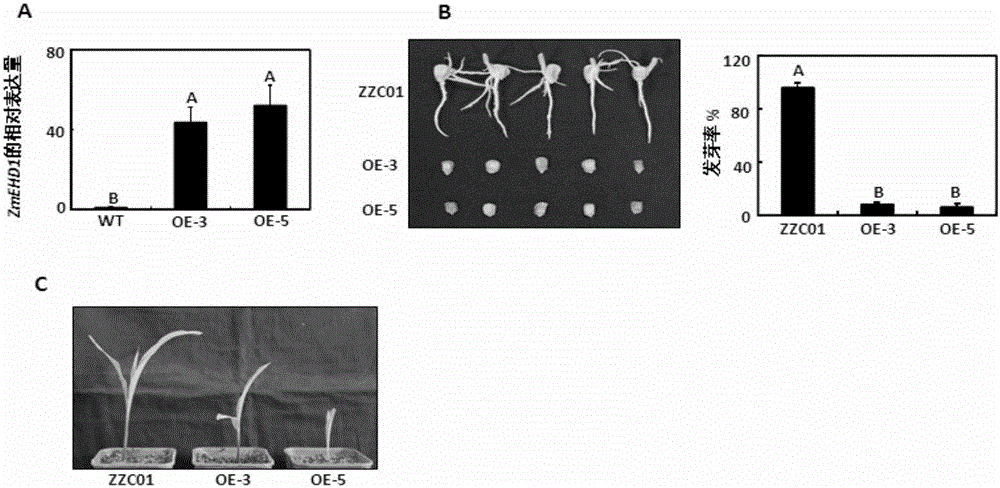
:內吞作用指質膜蛋白的內在轉化或者吸收胞外物質轉運到胞內的過程,分為依靠網絡蛋白介導的內吞作用(CME,clathrin-mediatedendocytosis)和不依靠網絡蛋白介導的內吞作用(Murphyetal.,2005)。在植物中,網絡蛋白介導的內吞作用是外界物質進入植物細胞的主要途徑(Fanetal.,2015)。與動物內吞作用的深入研究相比,最近幾年才證明網絡蛋白介導的內吞作用對植物發育的各個階段非常關鍵,包括決定細胞的極性(Vanetal.,2011;Wangetal.,2013),雄性生殖器官的發育(BlackbournandJackson,1996;Kimetal.,2013),配子的發育(Backuesetal.,2010)和胚胎的形成(Fanetal.,2013;Kitakuraetal.,2011)。與此同時,網絡蛋白介導的內吞作用在非生物和生物脅迫的植物響應方面也起著重要作用,這些脅迫是通過膜上的轉運體或者受體轉化成脅迫信號,例如硼轉運子(Takanoetal.,2010),鐵介導的轉運子(Barberonetal.,2011),生長素輸出載體(Dhonuksheetal.,2007),油菜素內酯受體(Dietal.,2013),細菌型鞭毛蛋白受體(Robatzeketal.,2006)和乙烯誘導木聚糖酶受體(BarandAvni,2009)。CME是一個復雜的過程,至少可以分為4個步驟:1,形成一個囊泡;2,將物質包裹在囊泡中;3,從質膜上釋放成熟的囊泡;4,與內體融合(Sigismundetal.,2012)。因為膜蛋白不能直接結合到質膜或者物質上,所以含有網絡蛋白內吞作用的囊泡的形成必須激活銜接蛋白復合體2(AP2)和質膜的聯系,因此AP2在網格蛋白介導的內吞作用方面起著重要作用。AP2復合體形成異源四聚體的復合物,包括兩個大亞基(β1-5和γ/α/δ/ε/ζ),一個中等亞基(σ1-5)和一個小亞基(σ1-5)(McMahonandBoucrot,2011)。除了典型的AP2復合體之外,其他的輔助蛋白也能把外界物質與質膜聯系起來,形成含有網格蛋白內吞作用的囊泡,例如C端Eps15同源結構域。玉米在主要的糧食作物中,總產量處于首位,并且也是生物能源和其他工業材料重要的原材料(Mclaren,2005)。作為生物學研究重要的模式系統,玉米在對植物發育和進化的認識中有著突出的貢獻,并且這些研究已經被用于闡釋玉米籽粒形態建成的發育機理。缺陷型籽粒(dek)突變體是最早在玉米中進行遺傳研究的材料之一。盡管幾百個缺陷型籽粒突變體基因已被分離,但是由于致死等位基因研究的難度,只有幾個相關的基因被分子克隆(ScanlonandTakacs,2009)。擬南芥ARF2通過抑制細胞分裂控制種子大小(Schruffetal.,2006),而水稻BIGGRAIN1(BG1)的活化通過調節生長素運輸顯著提高籽粒大小(Liuetal.,2015),同時,IAA-葡萄糖水解酶負調控水稻的胚乳發育(Ishimaruetal.,2013)。前人通過分析種子特異且可生長的玉米胚乳缺陷型突變體de18,闡明了玉米IAA合成關鍵基因ZmYUC1對于玉米正常的胚乳發育是必須的(Bernardietal.,2012)。因而,生長素的合成、轉運和信號可能協作調控玉米的營養發育和生殖發育(Gallavottietal.,2008;LiandLi,2016)。技術實現要素:本發明的一個目的是提供一種蛋白質。本發明提供的蛋白質是如下a)或b)或c)或d)的蛋白質:a)氨基酸序列是序列4所示的蛋白質;b)氨基酸序列是序列2所示的蛋白質;c)在序列4或序列2所示的蛋白質的N端和/或C端連接標簽得到的融合蛋白質;d)將序列4或序列2所示的氨基酸序列經過一個或幾個氨基酸殘基的取代和/或缺失和/或添加得到的具有相同功能的蛋白質。其中,序列2和序列4均由547個氨基酸殘基組成。為了使a)中的蛋白質便于純化,可在序列表中序列4或序列2所示的蛋白質的氨基末端或羧基末端連接上如表1所示的標簽。表1、標簽的序列標簽殘基序列Poly-Arg5-6(通常為5個)RRRRRPoly-His2-10(通常為6個)HHHHHHFLAG8DYKDDDDKStrep-tagII8WSHPQFEKc-myc10EQKLISEEDL上述c)中的蛋白質,所述一個或幾個氨基酸殘基的取代和/或缺失和/或添加為不超過10個氨基酸殘基的取代和/或缺失和/或添加。上述c)中的蛋白質可人工合成,也可先合成其編碼基因,再進行生物表達得到。上述c)中的蛋白質的編碼基因可通過將序列1或序列3所示的DNA序列中缺失一個或幾個氨基酸殘基的密碼子,和/或進行一個或幾個堿基對的錯義突變,和/或在其5′端和/或3′端連上表1所示的標簽的編碼序列得到。本發明的另一個目的是提供與上述蛋白質相關的生物材料。本發明提供的與上述蛋白質相關的生物材料為下述A1)至A12)中的任一種:A1)編碼上述蛋白質的核酸分子;A2)含有A1)所述核酸分子的表達盒;A3)含有A1)所述核酸分子的重組載體;A4)含有A2)所述表達盒的重組載體;A5)含有A1)所述核酸分子的重組微生物;A6)含有A2)所述表達盒的重組微生物;A7)含有A3)所述重組載體的重組微生物;A8)含有A4)所述重組載體的重組微生物;A9)含有A1)所述核酸分子的轉基因植物細胞系;A10)含有A2)所述表達盒的轉基因植物細胞系;A11)含有A3)所述重組載體的轉基因植物細胞系;A12)含有A4)所述重組載體的轉基因植物細胞系。上述相關生物材料中,A1)所述核酸分子為如下1)或2)或3)或4)所示的基因:1)其編碼序列是序列3所示的cDNA分子或DNA分子;2)其編碼序列是序列1所示的cDNA分子或DNA分子;3)與1)或2)限定的核苷酸序列具有75%或75%以上同一性,且編碼上述蛋白質的cDNA分子或基因組DNA分子;4)在嚴格條件下與1)或2)或3)限定的核苷酸序列雜交,且編碼上述蛋白質的cDNA分子或基因組DNA分子。其中,所述核酸分子可以是DNA,如cDNA、基因組DNA或重組DNA;所述核酸分子也可以是RNA,如mRNA或hnRNA等。本領域普通技術人員可以很容易地采用已知的方法,例如定向進化和點突變的方法,對本發明的編碼上述蛋白質的核苷酸序列進行突變。那些經過人工修飾的,具有編碼上述蛋白質的核苷酸序列75%或者更高同一性的核苷酸,只要編碼上述蛋白質且具有相同功能,均是衍生于本發明的核苷酸序列并且等同于本發明的序列。這里使用的術語“同一性”指與天然核酸序列的序列相似性。“同一性”包括與本發明的編碼序列2或序列4所示的氨基酸序列組成的蛋白質的核苷酸序列具有75%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或計算機軟件進行評價。使用計算機軟件,兩個或多個序列之間的同一性可以用百分比(%)表示,其可以用來評價相關序列之間的同一性。上述75%或75%以上同一性,可為80%、85%、90%或95%以上的同一性。本發明還有一個目的是提供上述蛋白質或上述相關生物材料的新用途。本發明提供了上述蛋白質或上述相關生物材料在調控植物體內生長素含量中的應用。上述應用中,所述調控為提高或降低。上述應用中,上述1)所述蛋白質或其相關生物材料是降低植物體內生長素含量。上述應用中,上述2)所述蛋白質或其相關生物材料是提高植物體內生長素含量。本發明還提供了上述蛋白質或上述相關生物材料在調控植物籽粒發育和/或營養生長中的應用。上述應用中,所述調控為促進或抑制。上述應用中,上述1)所述蛋白質或其相關生物材料是抑制植物籽粒發育和/或營養生長。上述應用中,上述2)所述蛋白質或其相關生物材料是促進植物籽粒發育和/或營養生長。本發明還提供了上述蛋白質或上述相關生物材料在調控植物內吞作用中的應用。上述應用中,所述調控植物內吞作用體現在與ZmAP2σ亞基互作影響植物網絡蛋白介導的內吞作用。上述應用中,所述影響為與ZmAP2σ亞基互作維持植物網絡蛋白介導的內吞作用,使植物網絡蛋白介導的內吞作用順利進行。本發明還有一個目的是提供上述1)所述的蛋白質或與1)所述的蛋白質相關的生物材料的新用途。本發明提供了上述1)所述的蛋白質或與1)所述的蛋白質相關的生物材料在培育生長素含量降低和/或籽粒發育延緩的轉基因植物中的應用。本發明的最后一個目的是提供一種培育生長素含量降低和/或籽粒發育延緩的轉基因植物的方法。本發明提供的培育生長素含量降低和/或籽粒發育延緩的轉基因植物的方法包括提高上述1)所述的蛋白質活性,得到轉基因植物的步驟;所述轉基因植物的生長素含量低于所述受體植物;所述轉基因植物的籽粒發育延緩于所述受體植物。上述方法中,所述轉基因植物的籽粒發育延緩于所述受體植物體現為如下a1)-a5)中的任一種:a1)所述轉基因植物的百粒重低于所述受體植物;a2)所述轉基因植物的發芽率低于所述受體植物;a3)所述轉基因植物的初生根短于所述受體植物;a4)所述轉基因植物的側根數少于所述受體植物;a5)所述轉基因植物的植株小于所述受體植物。上述方法中,所述提高上述1)所述的蛋白質活性的方法為將上述1)所述的蛋白質活性的編碼基因導入受體植物。上述方法中,上述1)所述的蛋白質活性的編碼基因的核苷酸序列為序列3。上述方法中,所述受體植物為單子葉植物或雙子葉植物,所述單子葉植物具體為玉米。本發明從玉米中克隆了ZmEHD1基因,并構建得到籽粒發育和營養生長受損的轉ZmEHD1mut玉米。通過實驗研究表明:和野生型玉米植株相比,轉ZmEHD1mut玉米的生長素含量降低,且外源施加有活性的NAA能夠提高轉ZmEHD1mut玉米的種子發芽率。ZmEHD1通過與ZmAP2σ亞基互作影響CME過程,且ZmEHD1介導的CME過程通過調節植物體內生長素含量而對玉米籽粒發育和營養生長起著至關重要的作用。附圖說明圖1為轉ZmEHD1mut玉米純合株系的表達量及發芽率檢測。其中,OE3和OE5均為轉ZmEHD1mut玉米純合株系;ZZC01為轉空載體植株;WT為野生型玉米植株。圖2為轉ZmEHD1mut玉米純合株系的籽粒表型。其中,WT為野生型玉米植株,OE為轉ZmEHD1mut玉米純合株系。圖3為轉ZmEHD1mut玉米純合株系和野生型玉米在授粉后15天、20天、25天和35天籽粒胚乳中ZmEHD1的表達情況。圖4為轉ZmEHD1mut玉米純合株系和野生型玉米的表型檢測。其中,OE為轉ZmEHD1mut玉米純合株系;WT為野生型玉米植株。圖5為轉ZmEHD1mut玉米純合株系的內吞作用受到抑制。其中,OE3和OE5均為轉ZmEHD1mut玉米純合株系;ZZC01為轉空載體植株;WT為野生型玉米植株。圖6為玉米ZmEHD1蛋白的亞細胞定位。圖7為玉米AP2σ能夠回補擬南芥ap2σ突變體的表型。圖8為玉米ZmEHD1蛋白與玉米AP2σ互作。圖9為轉ZmEHD1mut玉米純合株系和野生型玉米的皮爾遜相關系數。其中,WT為野生型玉米植株,OE為轉ZmEHD1mut玉米純合株系。圖10為通過RT-PCR驗證RNA-seq結果。其中,WT為野生型玉米植株,OE為轉ZmEHD1mut玉米純合株系。圖11為不同濃度1-NAA處理轉ZmEHD1mut玉米純合株系種子。其中,WT為野生型玉米植株,OE為轉ZmEHD1mut玉米純合株系。圖12為用不同濃度赤霉素處理轉ZmEHD1mut玉米純合株系和野生型玉米種子。其中,WT為野生型玉米植株,OE為轉ZmEHD1mut玉米純合株系。具體實施方式下述實施例中所使用的實驗方法如無特殊說明,均為常規方法。下述實施例中所用的材料、試劑等,如無特殊說明,均可從商業途徑得到。下述實施例中的定量試驗,均設置三次重復實驗,結果取平均值。下述實施例中的pSPYNE載體和pSPYCE載體均在文獻“Walteretal.,2004Visualizationofproteininteractionsinlivingplantcellsusingbimolecularfluorescencecomplementation.PlantJ.2004Nov;40(3):428-38.”中公開過,公眾可從河南農業大學獲得。下述實施例中涉及的試劑及培養基的配方:1)20%葡萄糖:稱取20g葡萄糖,加入100ml蒸餾水,溶解充分后過濾除菌。2)0.2%腺嘌呤鹽酸鹽:稱取0.4g固體粉末,加入200ml蒸餾水,溶解充分后過濾除菌。3)SD-Trp缺陷培養基:稱取0.67gYNB(無氨基酸)培養基、0.074gDO-Trp、2g瓊脂粉,加入90ml蒸餾水,121℃滅菌15min,超凈臺中加入10ml20%的無菌葡萄糖,鋪平板。4)SD-Leu缺陷培養基:稱取0.67gYNB(無氨基酸)培養基、0.069gDO-Leu、2g瓊脂粉,加入90ml蒸餾水,121℃滅菌15min,超凈臺中加入10ml20%的無菌葡萄糖,鋪平板。5)SD-Leu-Trp缺陷培養基(三缺培養基):稱取0.67gYNB(無氨基酸)培養基、0.064gDO-Leu-Trp、2g瓊脂粉,加入90ml蒸餾水,121℃滅菌15min,超凈臺中加入10ml20%的無菌葡萄糖,鋪平板。6)SD-Leu-Trp-His缺陷培養基(四缺培養基):稱取0.67gYNB(無氨基酸)培養基、0.062gDO-Leu-Trp-His、2g瓊脂粉,加入90ml蒸餾水,121℃滅菌15min,超凈臺中加入10ml20%的無菌葡萄糖,鋪平板。7)SD-Leu-Trp-His-Ade缺陷培養基:稱取0.67gYNB(無氨基酸)培養基、0.060gDO-Leu-Trp-His-Ade、2g瓊脂粉,加入90ml蒸餾水,121℃滅菌15min,超凈臺中加入10ml20%的無菌葡萄糖,鋪平板。8)10×TE緩沖液(pH7.5):10ml1MTris-HCl(pH7.5)、2ml0.5MEDTA(pH8.0),定容至100ml后,121℃滅菌15min。9)50%的PEG-4000:稱取50gPEG-4000溶于100ml蒸餾水后,121℃滅菌15min。下述實施例中的CPB載體在文獻“WenwenLiu,QingSun,KaiWang,QingguoDuandWen-XueLiNitrogenLimitationAdaptation(NLA)isinvolvedinsource-to-sinkremobilizationofnitratebymediatingthedegradationofNRT1.7inArabidopsisNewPhytologist(2016)doi:10.1111/nph.14396”中公開過,公眾可從河南農業大學獲得。下述實施例中的抑制劑E-64D是CaymanChemical的產品,產品目錄號為13533。下述實施例中的擬南芥ap2σ亞基功能缺失的突變體在文獻“LushengFan1,2,*,HuaiqingHao1,*,YiqunXue1,2,LiangZhang3,KaiSong1,2,ZhaojunDing4,MiguelA.Botella5,HaiyangWang6andJinxingLin1,DynamicanalysisofArabidopsisAP2σsubunitrevealsakeyroleinclathrin-mediatedendocytosisandplantdevelopment.Development.2013Sep;140(18):3826-37.doi:10.1242/dev.095711.Epub2013Aug7.”中公開過,公眾可從河南農業大學獲得。下述實施例中的pCAMBIA3301載體在文獻“LiJ,FuJ,ChenY,FanK,HeC,ZhangZ,LiL,LiuY,ZhengJ,RenD,WangG.TheU6Biogenesis-Like1PlaysanImportantRoleinMaizeKernelandSeedlingDevelopmentbyAffectingthe3'EndProcessingofU6snRNA.MolPlant.2016Nov4.pii:S1674-2052(16)30263-5.doi:10.1016/j.molp.2016.10.016.[Epubaheadofprint]”中公開過,公眾可從河南農業大學獲得。實施例1、ZmEHD1蛋白的獲得以野生型玉米(浚9058)的cDNA為模板,采用F:ATGGATATCGCGCGGGCGGC和R:TCACTCGTACGGGTTCCGCA引物進行PCR擴增,得到PCR擴增產物,并對PCR擴增產物進行測序分析。測序結果表明:PCR擴增得到大小為1644bp的條帶,其核苷酸序列如序列1所示,將序列1所示的基因命名為ZmEHD1基因,ZmEHD1基因編碼的蛋白質的氨基酸序列如序列表中序列2所示,將序列2所示的蛋白質命名為ZmEHD1蛋白。ZmEHD1蛋白含有EHD家族的典型結構:一個有兩個calcium-bindingEF-hands(15-39aa和49-84aa)組成的EHdomain,一個P-loop(GQYSTGKT),一個dynamin-typeguaninenucleotide-bindingdomain,一個coil-coildomain。實施例2、轉ZmEHD1mut玉米的制備及其表型檢測一、轉ZmEHD1mut玉米的制備1、表達載體的構建以BamHI(NewEnglandBiolabs,R3136L)為限制性酶切位點,利用In-Fusion(takara,639648)方法將序列3所示的ZmEHD1mut基因序列克隆到pCAMBIA3301載體上,得到重組質粒pCAMBIA3301-ZmEHD1mut。該質粒包含了一個由CaMV35S啟動子啟動的鏈霉菌屬吸水鏈霉菌草丁膦乙酰轉移酶基因(bar)。重組質粒pCAMBIA3301-ZmEHD1mut表達序列4所示的ZmEHD1mut蛋白。ZmEHD1mut基因序列為將序列1所示的ZmEHD1基因的第366位由G突變為A,且第58位由T突變為C,且第633位由G突變為C,且第1172位由T突變為C,且第1299位由A突變為G,且第1302位由C突變為T,且第1396位由A突變為G,且第1443位由C突變為G,且第1473位由C突變為T,且保持序列1其他序列不變后得到的序列。ZmEHD1mut蛋白的氨基酸序列為將序列2第391位由V突變為A,且第466位由M突變為V,且第481位由D突變為E,且保持序列2其他序列不變后后得到的序列。2、重組菌的制備將步驟1制備的重組質粒pCAMBIA3301-ZmEHD1mut通過電擊法轉入農桿菌EHA105(北京華越洋生物科技有限公司,GX0133-100),得到重組菌pCAMBIA3301-ZmEHD1mut/EHA105。將步驟1制備的pCAMBIA3301載體通過電擊法轉入農桿菌EHA105,得到重組菌pCAMBIA3301/EHA105。3、轉ZmEHD1mut玉米的獲得及鑒定用重組菌pCAMBIA3301-ZmEHD1mut/EHA105侵染野生型玉米自交系Qi319的幼胚,得到轉基因植株。轉基因植株的培養和篩選的具體步驟參照文獻“X.-Q.Huang·Z.-M.WeiHigh-frequencyplantregenerationthroughcallusinitiationfrommatureembryosofmaize(ZeaMaysL.)PlantCellRep(2004)22:793–800DOI10.1007/s00299-003-0748-9”中的方法。用重組菌pCAMBIA3301/EHA105侵染野生型玉米自交系Qi319的幼胚,得到轉空載體玉米(ZZCO1)。用熒光定量PCR方法對轉基因植株做進一步檢測,得到陽性轉ZmEHD1mut玉米純合株系。熒光定量檢測的具體步驟如下:提取轉基因植株和野生型玉米植株葉片的RNA,反轉錄為cDNA,用F:GATGACCTCCTCTGTGACCTGAA,R:AAGTTGTCTATGAGTCGCTGCTG引物進行熒光定量PCR檢測。在96孔板中配制如下反應體系:cDNA2.0μl、SYBRPremixExTaqTM(2×)10μl、50×DyeⅡ0.4μl、PrimerF/PrimerR0.5/0.5μl,ddH2O補足20μl。使用ABI7500RealTimePCRSystem熒光定量PCR儀進行熒光定量PCR反應。每個基因設置三個重復,每個基因設置一個陰性對照,按如下程序進行擴增反應:95℃預變性30s;95℃變性5s,60℃退火30s,72℃延伸34s,40個循環。每個循環在延伸階段采集信號。反應結束后導出Ct值,利用比較Ct值分析基因的表達水平,同時瓊脂糖凝膠電泳檢測擴增的專一性及其準確性。結果如圖1A所示。陽性轉ZmEHD1mut玉米純合株系OE3和OE5中的ZmEHD1基因表達量顯著高于野生型。選取陽性轉ZmEHD1mut玉米純合株系OE3和OE5,用于下述實驗。二、轉ZmEHD1mut玉米的表型分析1、授粉后籽粒發育情況檢測對轉ZmEHD1mut玉米純合株系OE3和OE5及野生型玉米植株(野生型玉米自交系Qi319)進行授粉,并檢測授粉后的籽粒發育情況。授粉的具體步驟如下:前期,將要授粉的植株的雌穗在吐絲之前套上雌穗袋,在雄穗穎殼打開露出花藥時,提早套上雄穗袋。授粉時,將植株彎曲,使雄穗處于身后防止串粉,最后,一只手取下雌穗上的紙袋,另一只手小心滴將花粉均勻地灑在花絲上,將紙袋套在雌穗上,用大頭針將其固定在雌穗上,寫好標記。授粉后14天之前陽性轉ZmEHD1mut玉米純合株系OE3和OE5與野生型籽粒相似,但授粉后16天野生型籽粒已明顯變為淡黃色,而轉ZmEHD1mut玉米純合株系籽粒仍為乳白色(圖2A)。成熟后,陽性轉ZmEHD1mut玉米純合株系OE3和OE5胚乳皺縮,胚小(圖2B),導致其百粒重顯著下降(圖2C)。為了研究轉ZmEHD1mut玉米純合株系和野生型籽粒的發育情況,用石蠟切片觀察授粉后15天的籽粒胚的縱切面。石蠟切片的具體制備步驟參照文獻“Yu-JunLiu,Zhi-HuiXiu,RobertMeeley,andBao-CaiTan,EmptyPericarp5EncodesaPentatricopeptideRepeatProteinThatIsRequiredforMitochondrialRNAEditingandSeedDevelopmentinMaize。ThePlantCell,Vol.25:868–883,March2013,www.plantcell.org2013AmericanSocietyofPlantBiologists.Allrightsreserved.”中的方法。石蠟切片觀察胚的縱切面可以看出:轉ZmEHD1mut玉米純合株系胚的發育比野生型粒胚的發育要延緩(圖2D),胚乳細胞的排列也較為疏松(圖2E)。2、授粉后表達量檢測按照步驟一的3中的熒光定量PCR方法檢測轉ZmEHD1mut玉米純合株系和野生型玉米在授粉后15天、20天、25天和35天籽粒胚乳中ZmEHD1的表達情況。結果如圖3所示。結果表明:ZmEHD1在轉ZmEHD1mut玉米純合株系中的表達量均高于野生型,且隨著籽粒發育時間的延長,表達量呈下降趨勢。3、發芽率檢測用水浸泡種子8小時催芽,浸泡過程中保持通氣。浸泡后將種子移至滅過菌的濾紙上,保持濾紙濕潤,在26℃黑暗條件下萌發。每天觀察種子發芽情況,統計發芽率。用紙培的方法測定發芽率發現轉ZmEHD1mut玉米純合株系的發芽率明顯低于野生型玉米植株,只有3%(圖4A),而且與野生型對照相比,轉ZmEHD1mut玉米純合株系初生根短,側根數少(圖4B),并且98%的轉ZmEHD1mut玉米純合株系到第二完全展開葉前死去(圖4C)。轉ZmEHD1mut玉米純合株系OE3和OE5的籽粒的發芽率也明顯低于轉空載體玉米(ZZCO1)(圖1B)。上述結果表明ZmEHD1對玉米的營養生長和籽粒發育至關重要。4、植株大小檢測當發芽的轉ZmEHD1mut玉米純合株系OE3和OE5、野生型玉米和轉空載體玉米(ZZCO1)種子長到一葉一心時移栽到裝好土的培養盆里,在玉米培養間培養,觀察植株生長狀態。ZmEHD1mut玉米純合株系OE3和OE5的種子可以生長到兩葉期,但明顯比轉空載體玉米(ZZCO1)小(圖1C)。野生型玉米種子的生長情況和轉空載體玉米(ZZCO1)種子的生長情況無顯著性差異。5、內吞作用FM4-64在水相中沒有熒光,只有插入脂質生物膜后才顯示較強的熒光,胞吞回收的囊泡因吞進細胞內部而被熒光標記,利用這種熒光探針能夠對細胞的胞吞過程進行有效標記。采用FM4-64標記實驗來驗證玉米ZmEHD1蛋白在玉米內吞作用中發揮作用。具體步驟如下:將野生型玉米植株、轉ZmEHD1mut玉米純合株系OE3和OE5和轉空載體玉米(ZZCO1)的根用5μMFM4-64(LifeTechnologies,F34653)標記10分鐘,然后觀察隨著時間的延伸,植株內吞作用的變化。結果如圖5所示:野生型玉米植株的根標記20分鐘后可觀察到約40%細胞(40/105)出現熒光斑點,而此時轉ZmEHD1mut玉米純合株系仍然觀察不到熒光斑點(0/120)。標記60分鐘后,野生型玉米植株有73%細胞(62/85)出現熒光斑點,轉ZmEHD1mut玉米純合株系卻只有37%的細胞(26/70)出現(圖5A)。轉空載體玉米(ZZCO1)的根系與野生型玉米植株出現的同樣的現象(圖5B),說明轉ZmEHD1mut玉米純合株系中內吞作用受到抑制,玉米ZmEHD1蛋白在內吞作用中起著重要作用。實施例3、ZmEHD1蛋白的亞細胞定位一、野生型玉米中ZmEHD1蛋白的亞細胞定位1、以BamHI(NewEnglandBiolabs,R3136L)為限制性酶切位點,利用In-Fusion(takara,639648)方法將序列1所示ZmEHD1基因的CDS全長克隆到CPB載體上,得到表達載體,該表達載體表達ZmEHD1蛋白和黃色熒光蛋白(YFP)的融合蛋白(YFP-ZmEHD1融合蛋白)。2、將步驟1的表達載體導入煙草表皮細胞中,在煙草表皮細胞中表達YFP-ZmEHD1融合蛋白,YFP信號主要在膜上(圖6A),與紅色熒光染料FM4-64結果一致。3、用抑制劑E-64D(影響核內體和液泡的融合)處理已經注射農桿菌的煙草,檢測ZmEHD1蛋白的運輸。結果如圖6所示。當用E-64D處理時,YFP-ZmEHD1在煙草表皮細胞聚集,而且與FM4-64信號共存,這個結果說明質膜的內在化(圖6A)。二、轉ZmEHD1mut玉米純合株系中ZmEHD1mut蛋白的亞細胞定位1、以BamHI(NewEnglandBiolabs,R3136L)為限制性酶切位點,利用In-Fusion(takara,639648)方法將ZmEHD1mut基因序列克隆到CPB載體上,得到表達載體,該表達載體表達ZmEHD1mut蛋白和黃色熒光蛋白(YFP)的融合蛋白(YFP-ZmEHD1mut融合蛋白)。2、將步驟1的表達載體導入煙草表皮細胞中,在煙草表皮細胞中表達YFP-ZmEHD1mut融合蛋白,YFP信號主要在膜上(圖6A),與紅色熒光染料FM4-64結果一致。3、用抑制劑E-64D(影響核內體和液泡的融合)處理已經注射農桿菌的煙草,檢測ZmEHD1mut蛋白的運輸。結果表明:當沒有用E-64D處理時,YFP-ZmEHD1mut蛋白信號在煙草表皮細胞聚集并與FM4-64信號共存(圖6B)。此現象表明在轉ZmEHD1mut玉米純合株系中核內體和液泡的融合受到損壞。實施例4、ZmEHD1與ZmAP2σ亞基互作為了驗證ZmEHD1是否通過與ZmAP2σ亞基互作影響網絡蛋白介導的內吞作用進行了如下實驗。一、ZmAP2σ影響網絡蛋白介導的內吞作用1、表達載體的構建將玉米AP2σ亞基(ZmAP2σ,GRMZM2G052713)的cDNA序列(序列5)通過LR反應(invitrogen,11791020)連接到pMDC32(NTCC典型培養物保藏中心BioVector質粒載體菌種細胞基因保藏中心,pMDC32),得到重組載體。2、重組菌的制備將重組載體導入GV3101菌(普如汀生物技術(北京)有限公司,Biovector610135)中,得到重組菌。3、轉ZmAP2σ擬南芥的獲得將含有重組載體的農桿菌通過Floral-dip法侵染擬南芥ap2σ亞基功能缺失的突變體中,將侵染過的擬南芥進行培養,得到T0代轉ZmAP2σ擬南芥種子。將T0代轉ZmAP2σ擬南芥種子表面消毒后,播種于1/2MS+hyg培養基上篩選,得到T1代轉ZmAP2σ擬南芥株系#1和#3,并采用引物AP2RT-1F:GAGTTCCGCACGCACAAAGT和AP2RT-1R:TTCTCAAGCTCGCCCATCCT對T1代轉ZmAP2σ擬南芥株系#1和#3進行RT-PCR檢測。RT-PCR檢測結果表明:ZmAP2σ在擬南芥突變體中表達(圖7A、圖7B),ZmAP2σ基因的轉入對擬南芥ap2σ亞基功能缺失突變體的表型進行了恢復(圖7C)。表明ZmAP2σ與AtAP2σ在網絡蛋白介導的內吞作用方面具有相似的功能,ZmAP2σ可影響網絡蛋白介導的內吞作用。二、熒光雙分子互補實驗1、表達載體的制備(1)nYFP-ZmAP2σ載體的構建以BamHI(NewEnglandBiolabs,R3136L)為限制性酶切位點,利用In-Fusion(takara,639648)方法將序列5所示的DNA片段克隆到pSPYNE載體上,且保持pSPYNE載體的其他序列不變,得到nYFP-ZmAP2σ載體;(2)cYFP-ZmEHD1載體的構建以BamHI為限制性酶切位點,利用In-Fusion方法將序列1所示的ZmEHD1基因序列克隆到pSPYCE載體上,且保持pSPYCE載體的其他序列不變,得到cYFP-ZmEHD1載體;(3)cYFP-ZmEHD1mut載體的構建以BamHI為限制性酶切位點,利用In-Fusion方法將序列3所示的ZmEHD1mut基因序列克隆到pSPYCE載體上,且保持pSPYCE載體的其他序列不變,得到cYFP-ZmEHD1mut載體。2、重組菌的制備分別將步驟1制備的表達載體轉入農桿菌EHA105(北京華越洋生物科技有限公司,GX0133-100)中,分別得到含有nYFP-ZmAP2σ載體的重組菌、含有cYFP-ZmEHD1載體的重組菌和含有cYFP-ZmEHD1mut載體的重組菌。將pSPYNE載體轉入農桿菌EHA105中,得到含有pSPYNE載體的重組菌。將pSPYCE載體轉入農桿菌EHA105中,得到含有pSPYCE載體的重組菌。3、煙草的注射及YFP信號檢測通過農桿菌轉化的方法,分別將目的基因注射到本氏煙草表皮細胞。具體步驟如下:(1)重組菌的重懸分別將含有pSPYNE載體的重組菌、含有pSPYCE載體的重組菌、含有nYFP-ZmAP2σ載體的重組菌、含有cYFP-ZmEHD1載體的重組菌和含有cYFP-ZmEHD1mut載體的重組菌、含有p19質粒的重組菌(addgene,MLCC13519)2000g離心10min后收集菌體,之后用重懸液(MgCl20.406g,MES0.426g,按照2/1000量加入AS(100mM母液),使得AS終濃度為0.2mM后,定容到200毫升,調節pH值至6.0左右)進行重懸,調節菌液OD600值至1.0,同時將含有p19質粒的農桿菌用重懸液調節OD600值至0.6。(2)重組菌的侵染將重懸后的含有nYFP-ZmAP2σ載體的重組菌、含有cYFP-ZmEHD1載體的重組菌和含有p19質粒的重組菌等體積混合,室溫下黑暗孵育2h后注射本氏煙草表皮細胞。將重懸后的含有nYFP-ZmAP2σ載體的重組菌、含有cYFP-ZmEHD1mut載體的重組菌和含有p19質粒的重組菌等體積混合,室溫下黑暗孵育2h后注射本氏煙草表皮細胞。將重懸后的含有pSPYNE載體的重組菌、含有cYFP-ZmEHD1載體的重組菌和含有p19質粒的重組菌等體積混合,室溫下黑暗孵育2h后注射本氏煙草表皮細胞。將重懸后的含有pSPYCE載體的重組菌、含有cYFP-ZmEHD1mut載體的重組菌和含有p19質粒的重組菌等體積混合,室溫下黑暗孵育2h后注射本氏煙草表皮細胞。(3)熒光信號檢測將步驟(2)獲得的注射重組菌的煙草正常培養3天后用LSM700激光共聚焦顯微鏡觀察熒光信號。當nYFP-ZmAP2σ和cYFP-ZmEHD1同時注射時,可以在膜上觀察到YFP信號(圖8A)。同樣的,當nYFP-ZmAP2σ和cYFP-ZmEHD1mut同時注射時,也可以觀察到YFP信號,但信號不僅在膜上,而是擴散到細胞質(圖8A)。說明ZmEHD1蛋白和ZmEHD1mut蛋白均能與ZmAP2σ亞基互作。三、酵母雙雜實驗1、質粒構建以Sfi-HF內切酶為限制性酶切位點,利用In-Fusion方法將序列1所示的ZmEHD1基因序列克隆到pPR3-NubG質粒上,且保持pPR3-NubG質粒的其他序列不變,得到pPR3-ZmEHD1質粒;以Sfi-HF內切酶為限制性酶切位點,利用In-Fusion方法將序列3所示的ZmEHD1mut基因序列克隆到pPR3-NubG質粒上,且保持pPR3-NubG質粒的其他序列不變,得到pPR3-ZmEHD1mut質粒;以Sfi-HF內切酶為限制性酶切位點,利用In-Fusion方法將序列5所示的DNA片段克隆到pBT3-STE質粒上,且保持pBT3-STE質粒的其他序列不變,得到pBT3-ZmAP2σ質粒。2、Bait建立(1)在YPDA固體培養基上挑選新鮮的NMY51克隆(在文獻“WenwenLiu,QingSun,KaiWang,QingguoDuandWen-XueLiNitrogenLimitationAdaptation(NLA)isinvolvedinsource-to-sinkremobilizationofnitratebymediatingthedegradationofNRT1.7inArabidopsisNewPhytologist(2016)doi:10.1111/nph.14396”中公開過),接菌至50mlYPDA液體培養基中,30℃,200rpm振蕩過夜培養。(2)培養OD546值至0.6-0.8。如果OD546超過1.0,則用新鮮培養基稀釋至0.2后繼續培養至OD546為0.6。(3)2500g離心5min,收集菌體,用2.5ml無菌水重懸,得到酵母感受態細胞。(4)將鮭魚精DNA(北京立虹東方生物科技中心,D1626)在99℃條件下變性5min,重復一次,得到變性后的鮭魚精DNA。在1.5mlEP管中準備PEG/LiAcmix:50%PEG-40001.2ml、1MLiAc180μl、變性后的鮭魚精DNA125μl。(5)準備1.5μgpBT3-ZmAP2σ質粒,加入300μl步驟(4)中的PEG/LiAcmix,渦旋混勻。(6)向(5)中加入100μl步驟(1)-(3)制備的酵母感受態細胞,充分渦旋混勻1min。(7)將上述混合物42℃孵育45min。(8)700g離心5min,收集菌體。(9)用100μl0.9%的無菌NaCl重懸菌體,取適量涂布在SD-Leu培養基上,30℃培養2-4天。(10)挑選陽性克隆(含有pBT3-ZmAP2σ質粒的酵母菌NMY51),PCR檢測(F:atgatccggttcatcctgctgcaga和R:tcattccagcttctcaagctcgccca)是否含有目的質粒。陽性菌落可用終濃度為25%的甘油保菌。3、Bait的自激活驗證(1)將步驟2中的陽性菌落接種到10mlSD-Leu液體培養基中,30℃振蕩過夜培養。(2)將過夜培養的菌體2500g離心10min,收集菌體。用YPDA液體培養基重懸后接種到50mlYPDA液體培養基中,加入培養基調整起始OD546約0.15-0.2。(3)30℃,220rpm搖菌3-5h,至OD546值為0.6-0.7。(4)準備500μl的鮭魚精DNA,99℃變性5min,重復一次,得到變性后的鮭魚精DNA。(5)在10ml無菌EP管中準備LiAc/TEmix8ml:10×TEbuffer0.88ml、1MLiAc0.88ml、ddH2O6.24ml。(6)在50ml無菌EP管中準備PEG/LiAcmix12ml:10×TEbuffer1.2ml、1MLiAc1.2ml、50%PEG-40009.6ml。(7)將(3)中的菌液700g離心5min,得到菌體。用無菌水重懸沉淀后700g離心5min,得到菌體。(8)用1mlLiAc/TEmix重懸沉淀,700g離心5min,去上清,取沉淀。(9)將步驟(8)獲得的沉淀用1.2mlLiAc/TEmix重懸得到感受態細胞。(10)將pPR3-NubG6μg、變性后的鮭魚精DNA100μl、步驟(9)中制備的感受態細胞600μl、PEG/LiAcmix2.5ml混合,得到混合物,并將混合物渦旋振蕩1min,混合均勻。(11)30℃孵育45min,每隔15min混勻一次。(12)42℃孵育20min。700g離心15min,去上清,收集沉淀。向沉淀中加入3mlYPDA重懸。30℃,150rpm離心90min,收集沉淀。(13)700g離心5min,收集沉淀,加入2ml0.9%NaCl,取600μl涂抹平板SD-Trp-Leu、SD-Trp-Leu-His及SD-Trp-Leu-His-Ade。30℃培養4-6天,觀察每種培養基上轉化細胞的生長狀況。若在三缺和四缺培養基上有一定量菌斑的形成,說明存在自激活現象,需要用不同濃度的3-AT進行背景抑制,具體判斷方法見該試劑盒說明書33頁,最終將7.5mM的3-AT作為正確濃度。4、Prey質粒的導入重復步驟3,分別將質粒pPR3-ZmEHD1及pPR3-ZmEHD1mut導入到含有pBT3-ZmAP2σ質粒的酵母菌NMY51,分別得到重組酵母菌(AP2σ-Cub+NubG-ZmEHD1)和重組酵母菌(AP2σ-Cub+NubG-ZmEHD1mut),并將最終菌體涂布在含有7.5mM3-AT的SD-Trp-Leu、SD-Trp-Leu-His及SD-Trp-Leu-His-Ade上,30℃培養4-6天,觀察每種培養基上轉化細胞的生長狀況。選取SD-Trp-Leu-His-Ade上正常生長的白斑,進行酵母菌的菌落PCR檢測。同時設置pTSU2-APPbait庫的建立和pNubG-Fe65的轉化作為對照。5、檢測β-半乳糖苷酶的活性將在SD-Trp-Leu-His-Ade培養基上正常生長的酵母菌接種到10mlSD-Trp-Leu液體培養基中,30℃振蕩培養過夜。2500g離心5min,收集菌體,用5ml0.9%NaCl重懸菌體,重復操作一次。測定OD546值。取100μl菌體,用0.9%NaCl分別稀釋10倍、100倍、1000倍。取相同體積的不同濃度的菌體分別滴在SD-Trp-Leu-His-Ade、SD-Trp-Leu-Ade、SD-Trp-Leu-Ade+X-gal培養基上,30℃培養2-4天,觀察菌體生長狀況。結果如圖8B所示。ZmEHD1和ZmEHD1mut均能與AP2σ亞基互作且互作的結果在缺His和添加β半乳糖苷酶活性的培養基上進行了驗證。說明ZmEHD1和ZmEHD1mut均可與ZmAP2σ亞基互作影響植物網絡蛋白介導的內吞作用。實施例5、ZmEHD1在調控植物體內生長素平衡中的應用一、轉ZmEHD1mut玉米純合株系的轉錄組分析為了深入探究參與ZmEHD1介導信號途徑的分子機制,利用RNA-seq技術比對分析轉ZmEHD1mut玉米純合株系(OE)的轉錄組分析和野生型玉米植株(WT)15天胚乳的完整轉錄組。每個樣本進行兩個生物學重復,通過Illumina高通量測序技術進行測序建庫。4個RNA庫總共獲得了超過5億個原始reads,通過過濾去除接頭污染reads、高含N比reads和低質量reads后,大約97%的reads得到保留。其中,超過1.2億的reads能夠完全與玉米B73RefGen_V3.27參考序列(ftp://ftp.ensemblgenomes.org/pub/plants/release-27/fasta/zea_mays)匹配。把不能匹配到玉米基因組的序列刪除,只有完全匹配的序列進行深入的分析。以每百萬reads中map到外顯子每Kb的reads個數(RPKM)來檢測每個基因的豐度。計算了每個處理的兩個生物學重復的相關系數(R值),考察重復之間的變異性。兩個處理重復間的R值都達到0.99(圖9),表明兩個生物學重復高度相關。依據Log2≥1并且顯著值≤0.05的條件,利用DEGseq軟件鑒定了4760個基因為差異表達基因(DEGs),其中包括ZmEHD1。對轉ZmEHD1mut玉米純合株系而言,這些差異基因中有2208個基因上調表達及2551個基因下調表達。通過實時熒光定量PCR來確定RNA-seq的結果。GRMZM2G088273、GRMZM2G346897、GRMZM2G067929和GRMZM2G418119表達水平與RNA-seq數據一致,在轉ZmEHD1mut玉米純合株系中表達量低于野生型(圖10A)。如預期的一樣,轉ZmEHD1mut玉米純合株系中GRMZM2G156877和GRMZM2G420988的表達量高于野生型(圖10A),表明了RNA-seq數據真實可靠。然后進行了基因本體分析(GO;http://bioinfo.cau.edu.cn/agriGO/),確定與差異表達基因相關的分子機制。GO分析結果表明4760個DEGs高度富集于幾個生物學過程,包括應激反應(GO:0006950,P=1.2e-12)、integraltomembrane(GO:0016021,P=1e-5)、intracellularmembrane-boundedorganelle(GO:0043231,P=0.0002)、液泡(GO:0005773,P=0.0007)、淀粉代謝過程(GO:0005982,P=1.6e-5)和糖類代謝過程(GO:0006950,P=1.2e-12)。一些DEGs具有已知或被推測的參與生長素介導信號途徑(GO:0009734,P=1.2e-25)和生長素刺激響應(GO:0009733,P=2.1e-25)的功能,其中包括生長素響應因子、AUX/IAA轉錄因子、生長素上調小RNA(SAUR)、吲哚乙醛氧化酶、生長素運輸蛋白和輸出載體(表1)。隨機選擇表1中的四個DEGs(GRMZM2G082943、GRMZM2G365188、GRMZM5G809195和GRMZM2G019799),通過實時熒光定量驗證了他們在轉ZmEHD1mut玉米純合株系和野生型中的不同表達水平(圖10B)。這些結果表明ZmEHD1可能影響玉米中生長素平衡。表1涉及植物生長素過程的差異表達基因二、ZmEHD1在調控植物體內生長素平衡中的應用1、野生型和轉ZmEHD1mut玉米純合株系的IAA含量檢測為了驗證ZmEHD1是否影響玉米生長素體內平衡,測定了野生型玉米植株(野生型玉米自交系Qi319)(WT)和轉ZmEHD1mut玉米純合株系OE3(OE)授粉后15天籽粒的IAA含量。IAA含量測定的具體步驟參照文獻“Wang,B.,Chu,J.,Yu,T.,Xu,Q.,Sun,X.,Yuan,J.,Xiong,G.,Wang,G.,Wang,Y.,andLi,J.(2015)Tryptophan-independentauxinbiosynthesiscontributestoearlyembryogenesisinArabidopsis.ProcNatlAcadSciUSA112:4821-4826.”中的方法。結果表明:轉ZmEHD1mut玉米純合株系(OE)籽粒中的IAA含量為2,558pgmg-1FW,和野生型玉米植株(WT)相比,轉ZmEHD1mut玉米純合株系OE3籽粒中的IAA含量降低了約30%(圖11A)。說明ZmEHD1蛋白可以調控植物體內生長素的含量。2、外源施加1-NAA可以提高轉ZmEHD1mut玉米純合株系的發芽率通過外源施加有活性的生長素化合物(1-NAA,1-naphthaleneaceticacid)來進一步驗證轉ZmEHD1mut玉米純合株系發芽率低是由生長素體內平衡失調造成。具體步驟如下:分別用如下濃度:5mg/L、10mg/L、20mg/L、30mg/L的1-NAA(PhytoTechnologyLaboratories,N600)水溶液和如下濃度:10mg/L、20mg/L、30mg/L、50mg/L的GA3(SIGMA-ALORICH,G7645)水溶液浸泡轉ZmEHD1mut玉米純合株系OE3種子12個小時,然后紙培(參照實施例2中的紙培方法),培養4天后,檢測種子發芽率。同時以水作為對照。30mgL-11-NAA水溶液和水均沒有提高轉ZmEHD1mut玉米純合株系種子的發芽率(圖11B,C)。但是當施加低濃度的1-NAA時,轉ZmEHD1mut玉米純合株系OE發芽率明顯提高,其中,施加10mgL-11-NAA最為明顯(圖11B)。同時,外源施加低濃度的1-NAA能使轉ZmEHD1mut玉米純合株系長到兩片完全展開葉時期。而外源施加GA3并不能提高轉ZmEHD1mut玉米純合株系的發芽率(圖12)。上述結果表明:外源施加1-NAA可以提高轉ZmEHD1mut玉米純合株系的發芽率。轉ZmEHD1mut玉米純合株系生長缺陷表型是由生長素體內平衡喪失引起的,說明ZmEHD1蛋白具有調控植物體內生長素平衡的功能。序列表<110>河南農業大學<120>一種調控植物體內生長素平衡的EHD1蛋白及其應用<160>5<210>1<211>1644bp<212>DNA<213>人工序列<220><223><400>1atggatatcgcgcgggcggctgctggcgtggcctgctgcaaggagcaccagaggatctac60gccgagtggttcgccctggccgacccagatggcgacggccgcgtcacgggcgccgatgcc120accagcttcttcgccatgtccgcgctttcacgcgccgatctcaagcaggtgtgggcgatc180gcggattccaggcggcaggggtacctcggattcgccgagttcgtcacagcaatgcagctc240gtgtctctggcccaagcagggaacgagatcacacaggacagtctgaaacgtgaagacctg300agcaccttggatccgcctgtgatggaaggtgttgatgagctactggctagatccaaggct360gttgtgaagagagttcacccagatgacaatggcaccactcaggcccaagcaccttccata420tatcactggtttggttcaaaatcagcacagaaggtgcagatgcctctgactgctgttacc480tctgtaattgatggcttgaaaagactctatgttgaaaaactgaagcctttggaagtcgcc540tacagatacaatgatttttctcacccattattgacaaacagtgattttgaagcaaagcca600atggttatgctcttgggtcagtattcaacagggaaaacaacattcattaagcatctgcta660aagacaagctacccaggagctcacgtcggaccagagccaacaactgatagattcgtagtt720gtcatgtcaggatctgatggaaggaccattcctggcaatactattgccgttcaagccgac780atgccattcactggtcttacaacctttgggggagcgtttctgtcgaagtttgaatgctct840cagatgccacatcccctgctagagcatatcacctttgtggacactcctggtgttctctct900ggggaaaagcaacggacccaacgtagctatgatttcactggtgtgacctcatggtttgct960gccaagtgtgaccttatccttcttctgtttgaccctcacaagcttgatataagtgatgag1020ttcaaacgtgtcatttcatctttacgtgggcatgatgacaaaatacgcgtagtgttgaac1080aaggccgaccaagttgacacacaacagcttatgagagtgtatggtgcattgatgtggtct1140cttgggaaagtgctaaatactcctgaggttgtgcgtgtgtatatcggatcatttaatgat1200aagccagtgaatgattcagctgttgggccaattggaaaagacttgtttgagagggagcaa1260gatgacctcctctgtgacctgaaggacattcctaagaaagcctgtgatcgacgggtcaat1320gagtttgttaagcgtgcccgagctgccaagatccatgcttacataattggccatctgaag1380aaggagatgcctgcaatgatgggaaaagccaaggctcagcagcgactcatagacaactta1440gacgaggagtttgcaaaggtgcagagggaataccatctccccgccggagactttcccgac1500gtcgagcacttcaagcaggtgctgggcgggtacagcatcgacaagttcgagaagctgaag1560ccgaagatggtgcaggcggtggacgacatgctcgctcacgacatcccggagctcctcaag1620aacttgcggaacccgtacgagtga1644<210>2<211>547<212>PRT<213>人工序列<220><223><400>2MetAspIleAlaArgAlaAlaAlaGlyValAlaCysCysLysGluHis151015GlnArgIleTyrAlaGluTrpPheAlaLeuAlaAspProAspGlyAsp202530GlyArgValThrGlyAlaAspAlaThrSerPhePheAlaMetSerAla354045LeuSerArgAlaAspLeuLysGlnValTrpAlaIleAlaAspSerArg505560ArgGlnGlyTyrLeuGlyPheAlaGluPheValThrAlaMetGlnLeu65707580ValSerLeuAlaGlnAlaGlyAsnGluIleThrGlnAspSerLeuLys859095ArgGluAspLeuSerThrLeuAspProProValMetGluGlyValAsp100105110GluLeuLeuAlaArgSerLysAlaValValLysArgValHisProAsp115120125AspAsnGlyThrThrGlnAlaGlnAlaProSerIleTyrHisTrpPhe130135140GlySerLysSerAlaGlnLysValGlnMetProLeuThrAlaValThr145150155160SerValIleAspGlyLeuLysArgLeuTyrValGluLysLeuLysPro165170175LeuGluValAlaTyrArgTyrAsnAspPheSerHisProLeuLeuThr180185190AsnSerAspPheGluAlaLysProMetValMetLeuLeuGlyGlnTyr195200205SerThrGlyLysThrThrPheIleLysHisLeuLeuLysThrSerTyr210215220ProGlyAlaHisValGlyProGluProThrThrAspArgPheValVal225230235240ValMetSerGlySerAspGlyArgThrIleProGlyAsnThrIleAla245250255ValGlnAlaAspMetProPheThrGlyLeuThrThrPheGlyGlyAla260265270PheLeuSerLysPheGluCysSerGlnMetProHisProLeuLeuGlu275280285HisIleThrPheValAspThrProGlyValLeuSerGlyGluLysGln290295300ArgThrGlnArgSerTyrAspPheThrGlyValThrSerTrpPheAla305310315320AlaLysCysAspLeuIleLeuLeuLeuPheAspProHisLysLeuAsp325330335IleSerAspGluPheLysArgValIleSerSerLeuArgGlyHisAsp340345350AspLysIleArgValValLeuAsnLysAlaAspGlnValAspThrGln355360365GlnLeuMetArgValTyrGlyAlaLeuMetTrpSerLeuGlyLysVal370375380LeuAsnThrProGluValValArgValTyrIleGlySerPheAsnAsp385390395400LysProValAsnAspSerAlaValGlyProIleGlyLysAspLeuPhe405410415GluArgGluGlnAspAspLeuLeuCysAspLeuLysAspIleProLys420425430LysAlaCysAspArgArgValAsnGluPheValLysArgAlaArgAla435440445AlaLysIleHisAlaTyrIleIleGlyHisLeuLysLysGluMetPro450455460AlaMetMetGlyLysAlaLysAlaGlnGlnArgLeuIleAspAsnLeu465470475480AspGluGluPheAlaLysValGlnArgGluTyrHisLeuProAlaGly485490495AspPheProAspValGluHisPheLysGlnValLeuGlyGlyTyrSer500505510IleAspLysPheGluLysLeuLysProLysMetValGlnAlaValAsp515520525AspMetLeuAlaHisAspIleProGluLeuLeuLysAsnLeuArgAsn530535540ProTyrGlu545<210>3<211>1644bp<212>DNA<213>人工序列<220><223><400>3atggatatcgcgcgggcggctgctggcgtggcctgctgcaaggagcaccagaggatctac60gccgagtggttcgccctggccgacccagatggcgacggccgcgtcacgggcgccgatgcc120accagcttcttcgccatgtccgcgctttcacgcgccgatctcaagcaggtgtgggcgatc180gcggattccaggcggcaggggtacctcggattcgccgagttcgtcacagcaatgcagctc240gtgtctctggcccaagcagggaacgagatcacacaggacagtctgaaacgtgaagacctg300agcaccttggatccgcctgtgatggaaggtgttgatgagctactggctagatccaaggct360gttgtaaagagagttcacccagatgacaatggcaccactcaggcccaagcaccttccata420tatcactggtttggttcaaaatcagcacagaaggtgcagatgcctctgactgctgttacc480tctgtaattgatggcttgaaaagactctatgttgaaaaactgaagcctttggaagtcgcc540tacagatacaatgatttttctcacccattattgacaaacagtgactttgaagcaaagcca600atggttatgctcttgggtcagtattcaacaggcaaaacaacattcattaagcatctgcta660aagacaagctacccaggagctcacgtcggaccagagccaacaactgatagattcgtagtt720gtcatgtcaggatctgatggaaggaccattcctggcaatactattgccgttcaagccgac780atgccattcactggtcttacaacctttgggggagcgtttctgtcgaagtttgaatgctct840cagatgccacatcccctgctagagcatatcacctttgtggacactcctggtgttctctct900ggggaaaagcaacggacccaacgtagctatgatttcactggtgtgacctcatggtttgct960gccaagtgtgaccttatccttcttctgtttgaccctcacaagcttgatataagtgatgag1020ttcaaacgtgtcatttcatctttacgtgggcatgatgacaaaatacgcgtagtgttgaac1080aaggccgaccaagttgacacacaacagcttatgagagtgtatggtgcattgatgtggtct1140cttgggaaagtgctaaatactcctgaggttgcgcgtgtgtatatcggatcatttaatgat1200aagccagtgaatgattcagctgttgggccaattggaaaagacttgtttgagagggagcaa1260gatgacctcctctgtgacctgaaggacattcctaagaaggcttgtgatcgacgggtcaat1320gagtttgttaagcgtgcccgagctgccaagatccatgcttacataattggccatctgaag1380aaggagatgcctgcagtgatgggaaaagccaaggctcagcagcgactcatagacaactta1440gaggaggagtttgcaaaggtgcagagggaatatcatctccccgccggagacttccccgac1500gtcgagcacttcaagcaggtgctgggcgggtacagcatcgacaagttcgagaagctgaag1560ccgaagatggtgcaggcggtggacgacatgctcgctcacgacatcccggagctcctcaag1620aacttgcggaacccgtacgagtga1644<210>4<211>547<212>PRT<213>人工序列<220><223><400>4MetAspIleAlaArgAlaAlaAlaGlyValAlaCysCysLysGluHis151015GlnArgIleTyrAlaGluTrpPheAlaLeuAlaAspProAspGlyAsp202530GlyArgValThrGlyAlaAspAlaThrSerPhePheAlaMetSerAla354045LeuSerArgAlaAspLeuLysGlnValTrpAlaIleAlaAspSerArg505560ArgGlnGlyTyrLeuGlyPheAlaGluPheValThrAlaMetGlnLeu65707580ValSerLeuAlaGlnAlaGlyAsnGluIleThrGlnAspSerLeuLys859095ArgGluAspLeuSerThrLeuAspProProValMetGluGlyValAsp100105110GluLeuLeuAlaArgSerLysAlaValValLysArgValHisProAsp115120125AspAsnGlyThrThrGlnAlaGlnAlaProSerIleTyrHisTrpPhe130135140GlySerLysSerAlaGlnLysValGlnMetProLeuThrAlaValThr145150155160SerValIleAspGlyLeuLysArgLeuTyrValGluLysLeuLysPro165170175LeuGluValAlaTyrArgTyrAsnAspPheSerHisProLeuLeuThr180185190AsnSerAspPheGluAlaLysProMetValMetLeuLeuGlyGlnTyr195200205SerThrGlyLysThrThrPheIleLysHisLeuLeuLysThrSerTyr210215220ProGlyAlaHisValGlyProGluProThrThrAspArgPheValVal225230235240ValMetSerGlySerAspGlyArgThrIleProGlyAsnThrIleAla245250255ValGlnAlaAspMetProPheThrGlyLeuThrThrPheGlyGlyAla260265270PheLeuSerLysPheGluCysSerGlnMetProHisProLeuLeuGlu275280285HisIleThrPheValAspThrProGlyValLeuSerGlyGluLysGln290295300ArgThrGlnArgSerTyrAspPheThrGlyValThrSerTrpPheAla305310315320AlaLysCysAspLeuIleLeuLeuLeuPheAspProHisLysLeuAsp325330335IleSerAspGluPheLysArgValIleSerSerLeuArgGlyHisAsp340345350AspLysIleArgValValLeuAsnLysAlaAspGlnValAspThrGln355360365GlnLeuMetArgValTyrGlyAlaLeuMetTrpSerLeuGlyLysVal370375380LeuAsnThrProGluValAlaArgValTyrIleGlySerPheAsnAsp385390395400LysProValAsnAspSerAlaValGlyProIleGlyLysAspLeuPhe405410415GluArgGluGlnAspAspLeuLeuCysAspLeuLysAspIleProLys420425430LysAlaCysAspArgArgValAsnGluPheValLysArgAlaArgAla435440445AlaLysIleHisAlaTyrIleIleGlyHisLeuLysLysGluMetPro450455460AlaValMetGlyLysAlaLysAlaGlnGlnArgLeuIleAspAsnLeu465470475480GluGluGluPheAlaLysValGlnArgGluTyrHisLeuProAlaGly485490495AspPheProAspValGluHisPheLysGlnValLeuGlyGlyTyrSer500505510IleAspLysPheGluLysLeuLysProLysMetValGlnAlaValAsp515520525AspMetLeuAlaHisAspIleProGluLeuLeuLysAsnLeuArgAsn530535540ProTyrGlu545<210>5<211>429bp<212>DNA<213>人工序列<220><223><400>5atgatccggttcatcctgctgcagaaccggcaggggaaaacacggctggccaagtactat60gtcgctctcgaggactccgagaagcacaaggttgagtacgaggtgcatcggctcgtggtc120aaccgggaccccaagttcaccaacttcgtcgagttccgcacgcacaaagttatatatagg180agatatgcaggactttttttctctatatgtgtggacatcaccgacaacgaattggcatac240ttggaatgtatccatctgtttgttgagatattggaccatttcttcagcaatgtgtgtgaa300cttgatttagtatttaatttccacaaggtttacttgatattggatgagttcattcttgct360ggagagcttcaagaaacaagcaaaagggcaattatagagaggatgggcgagcttgagaag420ctggaatga429當前第1頁1 2 3
相關技術
網友詢問留言
已有0條留言
- 還沒有人留言評論。精彩留言會獲得點贊!
1