生菜作為宿主在表達蛋白和/或多肽中的應用的制作方法
本發明涉及生物技術領域,特別涉及生菜作為宿主在表達蛋白和/或多肽中的應用。
背景技術:
植物作為藥物蛋白質的表達和生產系統,已經研究了近三十年。除了低成本和產量高等的優點外,基于植物的表達系統還降低了人和動物病原體從產生蛋白質的過程中傳播到人類的風險。此外,植物真核蛋白內膜表達系統和分泌途徑類似于哺乳動物細胞。植物表達系統可大量生產大分子量以及多亞基的藥物蛋白質,且大大優于原核生物表達系統,比如大腸桿菌表達系統。如需要翻譯后修飾的,糖基化的蛋白,以及需要組裝的單克隆抗體,其不能用原核系統實現。利用植物生產的藥用蛋白作為生物試劑已經商業化,或用作家禽的疫苗添加劑。2012年,美國食品和藥物管理局(fda)批準了用于治療遺傳疾病1型戈謝病的蛋白elelysotm(taliglucerasealfa),而這種蛋白是利用胡蘿卜生產的。在過去十年中,人們對藥物蛋白質的需求急劇增加,所以fda批準用于臨床試驗的植物藥用蛋白質的數量也在不斷增加。
綠色熒光蛋白最早是由下村修等人在1962年在一種學名aequoreavictoria的水母中發現。其基因所產生的蛋白質,在藍色波長范圍的光線激發下,會發出綠色螢光。這個發光的過程中還需要冷光蛋白質aequorin的幫助,且這個冷光蛋白質與鈣離子(ca2+)可產生交互作用。
蚯蚓在中國,日本和其他遠東國家已經被用作傳統藥物數千年。口服干蚯蚓粉被認為有利于血液循環系統。蚓激酶(lumbrokinase)是從不同種類的蚯蚓分離和純化的一組酶。這些酶被認為可用于治療與血栓形成相關的各種病癥的纖維蛋白溶解劑。好多蚓激酶基因已經克隆。遺傳技術的進步提供了生產重組蚓激酶的能力,并且已經用于臨床試驗以及用于口服健保產品。至今位置,genbank已經存儲了24個蚓激酶基因序列。在大腸桿菌,山羊乳腺,酵母巴斯德畢赤酵母和植物中已經成功地表達蚓激酶。由于其易于處理和快速生長,大腸桿菌表達系統具有以低成本高產量的生產蚓激酶的潛力。然而,在大腸桿菌中產生的重組lks多為包涵體。因此,復性過程是恢復和重建酶活性所必需的。由于原核細胞不能進行翻譯后修飾,大腸桿菌可能無法進行適當的折疊,加工和糖基化來表達真核蚓激酶蛋白。研究表面,植物已經成為除細菌,酵母或哺乳動物細胞生產藥物的方便和經濟的替代品。植物具有實現蛋白質穩定性和生物活性必需的翻譯后修飾所需的機制。植物中的蛋白質合成途徑也與動物細胞非常相似。利用植物系統生產藥物蛋白質的成本比使用哺乳動物細胞培養物和微生物發酵系統便宜得多。一般來說,利用植物系統生產重組人類藥物蛋白質及其衍生產品的成本僅為利用哺乳動物細胞培養系統的0.1%和微生物系統的2%-10%。更重要的是,植物不是人類病原體的宿主。因此,來自植物的重組蛋白質不太可能將致病劑傳播給人類。
植物中表達重組蛋白有兩種主要方法:開發穩定遺傳的轉基因植物品系或在植物中瞬間表達外源蛋白質。第一個策略是將編碼該蛋白質的dna克隆到表達載體中,并將其遞送至植物的細胞核或質體基因組中。外源dna在植物中的穩定整合使得該蛋白在后代的表達,而且可在野外或溫室大規模生產。然而,在植物中穩定表達蛋白耗費人力物力。而且篩選表達量高,遺傳穩定,農藝性狀好的純合子品系通常需要幾年時間。另外,關于生物安全問題,如基因漂移、花粉污染和病蟲害防治也會引起生物技術監管機構的關注。而第二種方法是瞬時表達外源蛋白。外源基因不直接整合到植物基因組中,只在宿主細胞中瞬時表達所需的蛋白質。與穩定轉化相比,瞬時表達在較短的時間內(一周內)可產生高含量的蛋白質。這種方法最大限度地減少了生物安全問題,因為處理過的植物或組織通常是在完全封閉的設施或容器中開發,不存在生物污染問題。
植物瞬時表達系統還可以用來生產重組蛋白以應對緊急情況。2014年,唯一用于有效抵抗埃博拉病毒爆發的抗體治療藥物,zmapptm,就是利用農桿菌滲透方法在煙草葉片中生產。zmapp的功效和安全性為推動植物制藥業的行業開辟了道路。目前,煙草是瞬間蛋白表達最常見的宿主植物,并且已經開發了各種載體和農桿菌滲透方法,用于在短時間內進行大規模生產。然而,煙草具有高纖維含量和潛在的有毒化合物,如生物堿尼古丁,顯著增加了下游純化過程中的成本,極大地阻礙了植物外源蛋白藥物的進一步發展。
現階段有利用轉基因植物生產疫苗,蛋白。但是轉基因植物生產過程長,大概兩年,而且轉基因植物存在基因污染的問題,而且植物中含有大量蛋白,淀粉,多糖等,對下游純化要求比較高。目前有利用煙草葉片來瞬間表達藥用蛋白的技術,周期大概7天。煙草含有尼古丁,煙堿,各種有毒酚類物質,造成下游純化工藝困難。
與煙葉系統相比,生菜中含有更少的酚類和有毒化合物,因此提供一種生菜系統表達醫用蛋白或多肽具有重要的現實意義。
技術實現要素:
有鑒于此,本發明提供生菜作為宿主在表達蛋白和/或多肽中的應用。本發明利用兩種蛋白綠色熒光蛋白(gfp)以及蚓激酶來證明在溫和的條件下生菜表達平臺可以用來生產其他外源多肽和蛋白質。
為了實現上述發明目的,本發明提供以下技術方案:
本發明提供了生菜作為宿主在表達蛋白和/或多肽中的應用。
在本發明的一些具體實施方案中,所述蛋白為醫用蛋白。
在本發明的一些具體實施方案中,所述醫用蛋白選自抗體、酶、生長因子或疫苗。
在本發明的一些具體實施方案中,所述蛋白為綠色熒光蛋白或蚓激酶。
本發明還提供了一種表達載體,包括雙元植物載體以及所述應用中的蛋白和/或多肽的核苷酸序列。
在本發明的一些具體實施方案中,所述蛋白為蚓激酶,所述表達載體的構建方法包括如下步驟:
本發明選擇了蚓激酶pi239作為候選基因,其從雙歧桿菌(l.bimastus)中克隆。pi239先前在大腸桿菌和酵母系統表達并被證明中具有優異的抗血栓形成特征。
步驟1:在蚓激酶核苷酸序列的5’末端分別加入xbal限制性酶切位點,在3’末端分別加入bamh1限制性酶切位點,克隆到puc57載體中,獲得puc-lum克隆載體;蚓激酶pi239cdna序列(genbank:af433650.1)如seqidno.1所示;蚓激酶蛋白pi239全序列的氨基酸序列如seqidno.2所示;成熟蚓激酶蛋白pi239序列的氨基酸序列如seqidno.3所示;加入酶切位點的pi239dna序列如seqidno.4所示;
步驟2:通過kpnl/sacl從所述puc-lum克隆載體中獲得基因片段pi239,克隆至雙元植物載體pcam35s,獲得表達載體p35-lum。
在本發明的一些具體實施方案中,所述蛋白為綠色熒光蛋白,所述表達載體為含有gfp的植物雙元載體pcambia1302。
本發明還提供了所述的表達載體在表達蛋白和/或多肽中的應用。
本發明還提供了一種生菜作為宿主表達蛋白和/或多肽的方法,將所述的表達載體轉化到農桿菌中,通過農桿菌介導真空滲透入生菜組織后,提取、分離蛋白質,獲得蛋白和/或多肽。作為優選,所述農桿菌選自根瘤農桿菌;更優選的,根瘤農桿菌為lba4404。
在本發明的一些具體實施方案中,所述農桿菌介導真空滲透包括如下步驟:
步驟1:抽真空25~45s;
步驟2:保持真空(-95kpa)壓力30~60s;
步驟3:釋放壓力使得滲透液滲入所述植物組織;
重復上述步驟2~3次,避光處理4d。
用于真空農桿菌滲透的煙草植物的生長時間通常為4至6周。而本發明消除了植物生長周期,大大節省了前期培育植物的時間。本發明利用生菜來瞬時表達在較短的時間內(4d)可產生高含量的蛋白質。這種方法最大限度地減少了生物安全問題,因為處理過的生菜組織通常是在完全封閉的設施或容器中開發,不存在生物污染問題。生菜基本不含有植物有毒物質,而且其本身纖維少,利于下游的蛋白純化。
本發明利用兩種蛋白綠色熒光蛋白(gfp)以及蚓激酶來證明在溫和的條件下生菜表達平臺可以用來生產其他外源多肽和蛋白質。
附圖說明
為了更清楚地說明本發明實施例或現有技術中的技術方案,下面將對實施例或現有技術描述中所需要使用的附圖作簡單地介紹。
圖1示植物雙元pcambia1302;
圖2(a)示生菜農桿菌真空滲透的方法演示;
圖2(b)示無土栽培生菜;
圖2(c)示倒轉浸入含有農桿菌的滲透液中整個系統密閉真空處理;
圖3(a)示農桿菌真空滲透4天后;
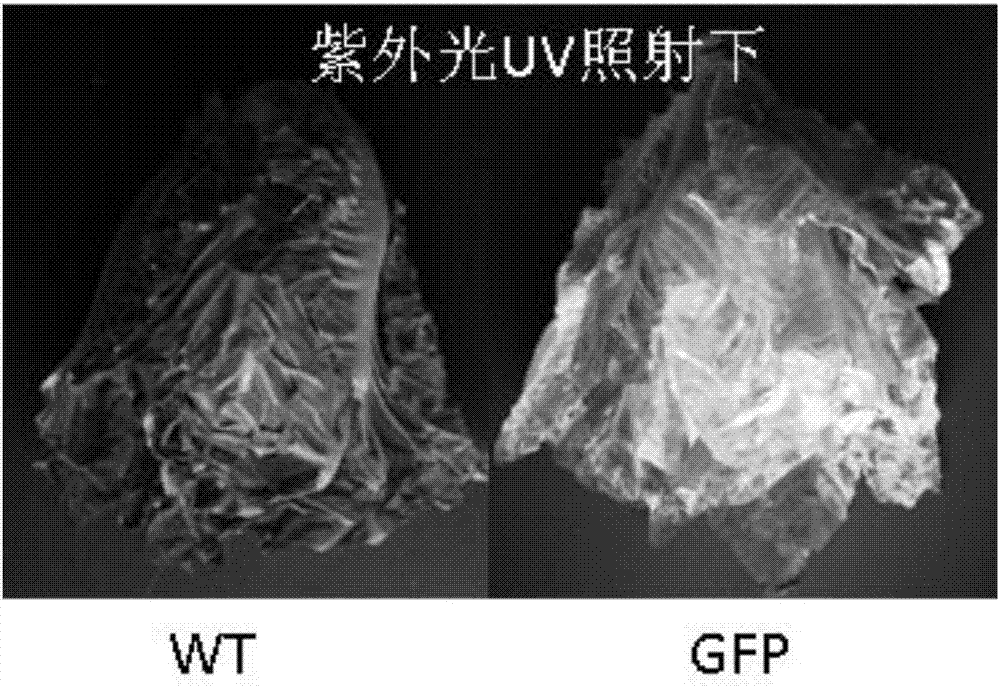
圖3(b)示在紫外光照射下,絕大多數生菜組織在真空滲透4d后顯示強烈的綠色熒光(gfp);相反,非農桿菌滲透生菜則在紫外光照射下顯示紅色(葉綠體);
圖4示粗提物(a)中gfp表達的可視化,最終純化(b);其中wt:非滲透生菜;gfp:農桿菌真空滲透生菜;
圖5示sds-page分析;泳道1:來自大腸桿菌的gfp;野生型生菜勻漿(泳道2,3)或純化后的gfp蛋白(4);箭頭顯示gfp(27kd)的大小
圖6示gfp濃度通過熒光光譜法測定gfp產量為0.37mg/g葉片重量;
圖7示植物瞬間表達載體示意圖;其中,35s,具有煙草花葉病毒(tmv)5‘utr的camv35s啟動子;nptii,用于卡那霉素抗性的編碼nptii基因的表達;nos3’,終止子;
圖8示sds-page(a)和western印跡(b)分析重組pi239;-ck:來自非滲透葉的陰性對照;
圖9示纖維蛋白平板測定;圖9(a)在纖維蛋白板上測試純化的洗脫液;圖9(b)裂解暈圈直徑;其中-ck:非滲透葉的洗脫(陰性對照);wt:pi239;lk:標準蚓激酶(18,000u/mg);值代表平均值±se;不同的字母表示顯著差異(p≤0.05;n=4;單因素方差分析測試);
圖10示體外人血液凝塊溶解試驗;圖10(a)重組pi239溶解人體血塊;圖10(b)血塊溶解百分比;pbs:1×磷酸鹽緩沖鹽水;-ck:非滲透假的洗脫(陰性對照);lk:標準蚓激酶(18,000u/mg);值代表平均值±se;不同的字母表示顯著差異(p≤0.01;n=3;單向方差分析測試)。
具體實施方式
本發明公開了生菜作為宿主在表達蛋白和/或多肽中的應用,本領域技術人員可以借鑒本文內容,適當改進工藝參數實現。特別需要指出的是,所有類似的替換和改動對本領域技術人員來說是顯而易見的,它們都被視為包括在本發明。本發明的方法及應用已經通過較佳實施例進行了描述,相關人員明顯能在不脫離本發明內容、精神和范圍內對本文所述的方法和應用進行改動或適當變更與組合,來實現和應用本發明技術。
本發明結果表明,生菜系統可以是更有效的表達平臺,為快速,瞬時表達重組蛋白質提供了方法。本文描述的真空農桿菌滲透方法簡單,快速,減少葉片壞死,而且可以提高重組蛋白產量。生菜可以通過承受真空壓力而增加蛋白質產量,并允許每片葉子更完整的滲透。由于生菜易于生長并且可商業上大量生產,因此比其他瞬時表達植物,如煙草等更容易獲得并且更便宜。并且由于不需要特殊設備或液氮,成本效益更高。本研究證明該方法可以用于短時間內大規模生產重組蛋白質。
本發明提供的生菜作為宿主在表達蛋白和/或多肽中的應用中所用原料及試劑均可由市場購得。
下面結合實施例,進一步闡述本發明:
實施例1利用生菜系統生產綠色熒光蛋白(gfp)
組織和根瘤土壤桿菌準備:
從當地蔬菜店購買了成熟生菜(紅葉,綠葉和長葉),放在標準生長臺燈下培養12h;或者將生菜種進行無土栽培如圖2(b)所示。將含有雙元植物載體pcambia1302(含有gfp,圖1)的根瘤農桿菌菌株lba4404接種到0.5lyeb(酵母提取物肉湯,5g/l蔗糖,5g/l胰蛋白胨,6g/l酵母提取物,0.24g/lmgso4,ph7.2)并補充抗生素液體培養基(50mg/l卡那霉素)。將接種的培養物在振蕩器(220rpm)中以25-28℃孵育72h。通過添加yeb培養基測量od600值并調節至3.5~4.5。然后收集培養液,離心(4500轉速)10min。將農桿菌細胞重懸在滲透培養基(10mmmes,10mmmgso4)中至o.d.600為0.5。
農桿菌介導的真空滲透:
優化了農桿菌真空滲透的方法(圖2)。將制備好的農桿菌培養懸浮液置于2l燒杯,并置于干燥器中。用刀將前10%的生菜切除,將其倒置(核心向上)并輕輕地旋轉于細菌懸浮液中,將干燥器密封,如圖2(c)所示。將真空泵(welchvacuum,niles,il,usa)打開以抽空,保持壓力狀態30~60s。然后打開該系統以釋放壓力,使滲透液滲入組織內的空間。該過程重復2~3次,直到清晰可見滲透液在生菜組織中有明顯擴散。然后將生菜組織從滲透液中輕輕取出,并用蒸餾水連續沖洗三次,然后轉移到塑料膜覆蓋的容器中。將處理的樣品在黑暗中保持4d,在uv光(395nm)下監測并拍照。實驗重復至少5次。
滲透后,絕大多數生菜組織在真空浸潤過程中淹沒,除了堅固的中肋區域外,其余部分均在真空滲透4d后顯示強烈的綠色熒光(圖3)。通過多次試驗觀察,該系統被證明非常有效。本發明優化了農桿菌浸潤,降低葉片壞死,增加蛋白質表達,提取,回收和純化過程。
蛋白質提取和分離:
經農桿菌真空滲透的生菜樣品用攪拌器攪拌,并用體積比為1:1比例的提取緩沖液(100mmkpi,ph7.8;5mmedta;10mmβ-巰基乙醇)攪拌機中高速勻漿1~2min。將勻漿物調節至ph為8.0,用紗布過濾,過濾物在4℃以10,000g離心15min以除去細胞碎片。收集上清液,與硫酸銨(50%)混合,并在冰上搖動孵育60min。通過離心機(10,000g)在4℃下再次分離15min。將得到的上清液進行第二輪硫酸銨(70%)沉淀,冰上搖動懸浮60min,再次在4℃下以10,000g離心15min。然后,棄去上清液,將處理樣品沉淀蛋白質溶于5ml緩沖液(20mmkpi,ph7.8;2mmedta;10mmβ-巰基乙醇)中并在4℃下儲存。gfp的產量通過分光熒光計(perkinelmerenspire2300multilabelreader,ma,usa)定量,發射峰在470~506nm之間。
植物來源的重組蛋白質的下游加工通常難于并且昂貴,因為纖維素細胞壁難以裂解以及次級植物代謝產物。攪拌機混合提取樣品中gfp的熒光肉眼可見(圖4)。用sds-page凝膠確認從真空滲透的生菜中檢測出重組gfp條帶(圖5)。此外,使用分光熒光分析(圖6)純化樣品的蛋白含量為0.37mg。
gfp可視化和sds-page分析:
為了證實gfp表達真實存在,收集粗制植物提取物和純化的最終蛋白質的樣品(5μl),并在黑暗的室內在uv光下顯現。將室溫或者熱變性(95℃)gfp加載緩沖液(biorad,hercules,ca,usa)在4~12%
通過熒光分光光度計進行gfp定量:
使用perkinelmerenspire2300multilabelreader來測量細菌對照gfp和生菜轉化的gfp,以比較次方法獲得的蛋白質量。使用來自商業樣品的已知量的gfp開發了標準曲線。通過對來自同種生菜(野生型wt)樣品的比較分析,從含有gfp的樣品中除去除背景蛋白熒光。將每個樣品的背景蛋白濃度的rfu繪制在標準曲線上,以確定每個樣品中gfp的近似量。使用從406-460nm之間的單個激發峰的發射數據計算相對熒光單位,發射峰在470-506nm之間。在這些參數內進行重復測量(n=12)。
實施例2利用生菜系統生產蚓激酶
植物表達構建體:
基于genbank(accessionno:af433650)的全長pi239dna序列,我們將限制性位點xbai位點連接到pi239的5'末端,在片段的3'末端加上bamh1位點,由genscriptinc(piscataway,nj,usa)合成,克隆到puc57載體中,得到克隆載體puc-lum。通過kpni/saci從puc-lum中分離出片段pi239,并克隆入植物雙元載體pcam35s(本實驗室保存),以生成植物表達載體p35-lum。隨后將植物表達載體p35-lum通過multiporator(eppendorf,hamburg,germany)電穿孔轉化到根瘤農桿菌ba4404中,用于隨后在生菜中的瞬時表達測定。利用sds-page凝膠電泳觀察純化重組蚓激酶蛋白的大小。
纖維蛋白溶解活性測定:
將瓊脂糖(0.2g)置于40ml的磷酸鹽緩沖鹽水(1xpbs)中,通過在微波中煮沸然后冷卻至約40℃。凝膠固化之前,將32mg人纖維蛋白原(sigma-aldrich,st.louis,mo,usa),10單位人纖溶酶原(rpeptidellc,bogart,ga,usa)和10單位人類凝血酶(biopharmlaboratoriesllc,bluffdale,ut,usa)加入到凝膠中并輕微旋轉混合以防止引入氣泡。然后將混合物倒入培養皿中并使其在室溫下固化。隨后在固化凝膠中制成孔(3mm直徑)。將10μl濃縮樣品(1μg/μl)稀釋在40μl的1xpbs緩沖液中,裝入凝膠孔中,并在室溫下孵育過夜。使用標準的蚓激酶(doctor'sbestinc,irvine,ca,usa)作為陽性對照(10μg),將來自非農桿菌滲透轉化的生菜葉片洗脫液用作陰性對照。測量清晰暈圈的實際直徑(mm)。與標準lk產品(18,000u/mg)相比,計算重組pi239的纖維蛋白溶解活性。平均值±se(n=4)通過單因素方差分析進行了分析。
本發明克隆了雙元植物表達載體p35-lum。在構建完成后,用特異性限制酶消化證實基因片段完整(圖7)。重組pi239經過sds-page分離(圖8a)。我們在泳道中觀察到估計分子量大約為37kda的條帶。在陰性對照泳道中沒有明顯的對應條帶。另外,通過hisprobe-hrp方法的western印跡分析也檢測到大約37kda的條帶(圖8b)。觀察到的分子量(37kda)大于pi239-his6鏈(32kda)的預測大小,這種差異最有可能是由于pi239蛋白的糖基化導致的。基于bradford測定法和sds-page上的條帶強度計算出得到的蛋白含量大概為135μg/g葉鮮重(lfw)。
血凝塊分析測定:
進行體外血液凝塊裂解測定。將購買的血塊(interstatebloodbank,memphis,tn,usa)切片成小片,并用1xpbs緩沖液徹底清洗。隨后,將血凝塊(約50mg/凝塊)轉移到24孔板中,并用1xpbs緩沖液沖洗兩次。將40μl濃縮樣品(1μg/μl)稀釋在460μl的1xpbs緩沖液中,并加入含有血塊的孔中。在37℃下孵育過夜后,測量未溶解的樣品,并在處理前后測定樣品的重量差異。pbs緩沖液和非轉染葉片的洗脫液用作陰性對照。標準蚓激酶(40μg)用作陽性對照。以重量差/未處理樣品的百分比計算凝塊裂解。平均值±se(n=3)通過單因素方差分析進行分析。
蚓激酶能夠通過直接溶解纖維蛋白來降解血塊。它還可以增強內源性t-pa活性,將纖溶酶原轉化為纖溶酶,導致纖維蛋白溶解。在本研究中,纖維蛋白平板測定顯示植物來源的蚓激酶具有明顯溶解纖維蛋白(圖9)。在商用蚓激酶和重組蛋白質的周圍顯示半透明的暈圈區域(圖9a)。重組蚓激酶pi239的裂解直徑分別為8.8mm,活性低于商業蚓激酶(18,000u/mg),其溶解直徑為12.25mm(圖9b)。重組pi239的相對纖維蛋白溶解活性計算為約13,400u/mg。植物來源的蚓激酶pi239體外可以溶解人血凝塊(圖10a)。通過凝塊自溶過程,在pbs孔和非滲透葉提取物中觀察到輕微裂解。當用標準蚓激酶和重組pi239處理凝塊時觀察到明顯的裂解。標準蚓激酶和重組蚓激酶分別顯示92%和76%的凝塊裂解(圖10b)。與標準蚓激酶相比,重組蚓激酶pi239的較低活性可能是由于來自純化過程中的植物成分或殘留洗脫液。在以前報道的研究中,最終洗脫液中存在的殘留植物污染物顯著干擾纖維溶解蛋白酶的酶活性。因此,只需要進一步優化純化策略獲得高純度的重組蚓激酶pi239,即可有效地增加血纖維蛋白溶解活性。
實施例3
對照組:利用煙葉生產綠色熒光蛋白和蚓激酶;
實驗組:本發明提供的生菜生產綠色熒光蛋白和蚓激酶;
表1綠色熒光蛋白和蚓激酶
*示與對照組相比p≤0.05;#示與對照組相比p≤0.01;
由表1可知,與對照組的煙葉系統相比,生菜瞬時表達綠色熒光蛋白和蚓激酶,顯著(p≤0.05)縮短了生產周期,顯著(p≤0.05)提高了蛋白含量,顯著(p≤0.05)提高了蛋白活性,簡化了蛋白純化的難易程度,極顯著(p≤0.01)降低了生產成本。
綜合上述試驗結果表明,植物系統尤其是生菜系統是更加經濟、高效的表達平臺。能夠快速瞬時表達重組蛋白質,可以在短時間內大規模生產綠色熒光蛋白和蚓激酶。
以上所述僅是本發明的優選實施方式,應當指出,對于本技術領域的普通技術人員來說,在不脫離本發明原理的前提下,還可以做出若干改進和潤飾,這些改進和潤飾也應視為本發明的保護范圍。
sequencelisting
<110>深圳惠升生物科技有限公司
<120>生菜作為宿主在表達蛋白和/或多肽中的應用
<130>mp1710876
<160>4
<170>patentinversion3.3
<210>1
<211>852
<212>dna
<213>蚓激酶pi239cdna序列
<400>1
atgttacttctcgctcttgcatcgttggtagcggtgggctttgcgcaaccaccagtctgg60
taccccggtggtcaatgcggtgtcagccagtactcagatgctggtgacatggaacttcct120
cccggaacaaaaattgtcggaggaattgaagccagaccatacgagttcccatggcaggtg180
tccgtccgaaggaagtcttccgattcccatttctgcggaggtagcatcatcaacgatcgt240
tgggttgtctgcgctgctcactgcatgcagggagagagccctgccctggtttcattggtc300
gtcggtgagcacgatagcagcgctgcgagtacagtacgtcagactcatgacgttgacagc360
atcttcgtccacgaggactacaacggaaatacctttgagaacgacgtttctgtcatcaag420
acagttaacgccatcgccatcgacatcaacgttgggccaatctgcgctccagatccagcc480
aacgattacgtctaccgtaagagccagtgctccggatggggaactatcaactcaggtgga540
gtctgctgccccaacgttctgcgatatgtgacactgaacgtcacaaccaacgccttctgc600
gatgatatctacagcccattatatacaattaccagcgacatgatctgcgccacggacaac660
accggacagaacgagagagactcttgccagggtgactctggcggccctctgagcgtcaag720
gatggcagcggaatcttcagcctcattggtattgtgtcttggggaatcggttgcgcatct780
ggatatccaggagtctacgcccgcgtcgggtcccaaactggatggatcacagacatcatc840
accaacaactaa852
<210>2
<211>283
<212>prt
<213>蚓激酶蛋白pi239全序列
<400>2
metleuleuleualaleualaserleuvalalavalglyphealagln
151015
proprovaltrptyrproglyglyglncysglyvalserglntyrser
202530
aspalaglyaspmetgluleuproproglythrlysilevalglygly
354045
ileglualaargprotyrglupheprotrpglnvalservalargarg
505560
lysserseraspserhisphecysglyglyserileileasnasparg
65707580
trpvalvalcysalaalahiscysmetglnglygluserproalaleu
859095
valserleuvalvalglygluhisaspserseralaalaserthrval
100105110
argglnthrhisaspvalaspserilephevalhisgluasptyrasn
115120125
glyasnthrphegluasnaspvalservalilelysthrvalasnala
130135140
ilealaileaspileasnvalglyproilecysalaproaspproala
145150155160
asnasptyrvaltyrarglysserglncysserglytrpglythrile
165170175
asnserglyglyvalcyscysproasnvalleuargtyrvalthrleu
180185190
asnvalthrthrasnalaphecysaspaspiletyrserproleutyr
195200205
thrilethrseraspmetilecysalathraspasnthrglyglnasn
210215220
gluargaspsercysglnglyaspserglyglyproleuservallys
225230235240
aspglyserglyilepheserleuileglyilevalsertrpglyile
245250255
glycysalaserglytyrproglyvaltyralaargvalglysergln
260265270
thrglytrpilethraspileilethrasnasn
275280
<210>3
<211>239
<212>prt
<213>成熟蚓激酶蛋白pi239序列
<400>3
ilevalglyglyileglualaargprotyrglupheprotrpglnval
151015
servalargarglysserseraspserhisphecysglyglyserile
202530
ileasnaspargtrpvalvalcysalaalahiscysmetglnglyglu
354045
serproalaleuvalserleuvalvalglygluhisaspserserala
505560
alaserthrvalargglnthrhisaspvalaspserilephevalhis
65707580
gluasptyrasnglyasnthrphegluasnaspvalservalilelys
859095
thrvalasnalailealaileaspileasnvalglyproilecysala
100105110
proaspproalaasnasptyrvaltyrarglysserglncyssergly
115120125
trpglythrileasnserglyglyvalcyscysproasnvalleuarg
130135140
tyrvalthrleuasnvalthrthrasnalaphecysaspaspiletyr
145150155160
serproleutyrthrilethrseraspmetilecysalathraspasn
165170175
thrglyglnasngluargaspsercysglnglyaspserglyglypro
180185190
leuservallysaspglyserglyilepheserleuileglyileval
195200205
sertrpglyileglycysalaserglytyrproglyvaltyralaarg
210215220
valglyserglnthrglytrpilethraspileilethrasnasn
225230235
<210>4
<211>866
<212>dna
<213>加入酶切位點的pi239dna序列
<400>4
tctagaatgttacttctcgctcttgcatcgttggtagcggtgggctttgcgcaaccacca60
gtctggtaccccggtggtcaatgcggtgtcagccagtactcagatgctggtgacatggaa120
cttcctcccggaacaaaaattgtcggaggaattgaagccagaccatacgagttcccatgg180
caggtgtccgtccgaaggaagtcttccgattcccatttctgcggaggtagcatcatcaac240
gatcgttgggttgtctgcgctgctcactgcatgcagggagagagccctgccctggtttca300
ttggtcgtcggtgagcacgatagcagcgctgcgagtacagtacgtcagactcatgacgtt360
gacagcatcttcgtccacgaggactacaacggaaatacctttgagaacgacgtttctgtc420
atcaagacagttaacgccatcgccatcgacatcaacgttgggccaatctgcgctccagat480
ccagccaacgattacgtctaccgtaagagccagtgctccggatggggaactatcaactca540
ggtggagtctgctgccccaacgttctgcgatatgtgacactgaacgtcacaaccaacgcc600
ttctgcgatgatatctacagcccattatatacaattaccagcgacatgatctgcgccacg660
gacaacaccggacagaacgagagagactcttgccagggtgactctggcggccctctgagc720
gtcaaggatggcagcggaatcttcagcctcattggtattgtgtcttggggaatcggttgc780
gcatctggatatccaggagtctacgcccgcgtcgggtcccaaactggatggatcacagac840
atctgagagctcttaattaaggatcc866
- 還沒有人留言評論。精彩留言會獲得點贊!