干細胞預防神經元頂梢枯死的用途的制作方法
本申請為分案申請,其原申請的申請日為2009年9月4日,申請號為200980144083.9,名稱為“干細胞預防神經元頂梢枯死的用途”。
本發明主要涉及神經元損傷的治療。具體地,本發明涉及減少因活化的巨噬細胞與營養不良的軸突相互作用而發生的軸突回縮(“頂梢枯死”),所述營養不良的軸突是在神經系統急性或慢性損傷過程中形成的。本發明還涉及促進軸突生長/再生。本發明特別涉及使用干細胞或其分泌的細胞因子,例如會在條件細胞培養基中產生的細胞因子,以改善或預防軸突頂梢枯死和/或促進軸突生長/再生。
背景技術:
軸突回縮
脊髓損傷后形成神經膠質疤痕,其造成cns再生的主要障礙(silverandmiller,2004)。在形成疤痕組織的區域中,再生軸突的末端延伸停止,并且膨脹并扭曲為可在軸突束(axontract)內保持數年的各種奇怪形狀的“生長錐”(ramonycajal,1928;liandraisman,1995;houleandjin,2001;kwonetal.,2002)。cns內的損傷軸突在初始損傷后的數小時到數周的時間內從軸索切斷的位置回縮。關于軸突回縮的性質、其原因、程度、時間以及對其是被動抑或主動過程的討論已有不同報道(fayazandtator,2000)。
體外神經膠質疤痕模型
在形成疤痕組織的區域內,多類生長抑制分子被上調,包括被稱為軟骨素/硫酸角質素蛋白聚糖(pg)的細胞外基質(ecm)分子的家族(fitchandsilver,1997;morgensternetal.,2002;jonesetal.,2003;tangetal.,2003)。pg大致呈梯度分布,在損傷邊緣濃度最低而在中心的濃度最高(daviesetal.,1999;fitchetal.,1999)。所述抑制性ecm組分阻斷有活性的神經膠質細胞供養軸突通過層粘連蛋白再生的可能性(mckeonetal.,1991)。微移植實驗顯示成體感覺神經元在遠離損傷時具有強再生能力。一旦再生纖維到達所述損傷位置附近,它們就能夠艱難進入損傷邊緣但最終在深入到最高pg濃度的區域時停止延伸并變得營養不良(daviesetal.,1999;grimpeandsilver,2004)。
在體外神經膠質疤痕模型中,使用尖緣的(即條紋)基底測定來檢查pg對軸突的作用,誘導了生長錐轉向或萎縮,但未發生營養不良(snowetal.,1990)。然而,在神經膠質疤痕的最新模型中,大致呈梯度的pg足以在再生成體軸突中產生營養不良的末端(tometal.,2004)。此體外系統迫使成體感覺神經元的再生軸突應對與層粘連蛋白混合的pg聚集蛋白聚糖的斑點梯度。在此神經膠質疤痕模型中形成了球狀多囊泡末端。pg導致生長錐營養不良和動態營養不良末端。
神經元組織中的炎癥和損傷
脊髓損傷的環境極端復雜。神經膠質疤痕的組分,例如高度硫酸化的蛋白聚糖、eph、開口和髓磷脂膜碎片(silverandmiller,2004;yiuandhe,2006;buschandsilver,2007),以及神經性炎癥過程均是導致再生障礙的原因。炎性細胞在損傷內積聚(fitchetal.,1999)。星形細胞離開損傷的中心,變大并上調抑制性硫酸軟骨素蛋白聚糖(cspg)的產生,cspg轉而導致在切斷的纖維上形成營養不良的末端球狀物(tometal.,2004)。損傷內的少突膠質細胞死亡,引起脫髓鞘,這導致高濃度的抑制性髓磷脂分解產物(yiuandhe,2006;xieandzheng,2008)。
盡管蛋白聚糖和髓磷脂對軸突生長的抑制性作用得到公認,然而神經性炎癥在再生和再生障礙中的作用仍然飽受爭議(popovichandlongbrake,2008)。然而,研究已經表明巨噬細胞滲入導致損傷大小增加,再生纖維的生長下降,以及幸免于初始損傷的神經元死亡增加(fitchetal.,1999;mcphailetal.,2004;donnellyandpopovich,2007)。活化的巨噬細胞和中性白細胞的負面影響被認為由細胞因子、類二十烷酸、自由基和蛋白酶的分泌介導,這些物質對神經元和神經膠質可能均有毒性(donnellyandpopovich,2007)。其中在脊髓損傷后巨噬細胞被清除、抑制或失活的多項研究已經報道了神經保護、再生增加以及運動、感覺和自主功能的改善(oudegaetal.,1999;popovichetal.,1999;mcphailetal.,2004;stirlingetal.,2004)。
技術實現要素:
本發明部分地基于發明人的以下發現:在體外神經膠質疤痕模型中,軸突回縮(頂梢枯死)ed-1+細胞例如活化的巨噬細胞和小神經膠質可通過外部給予某些類型的細胞或培養過所述細胞的條件細胞培養基而減少。這些體外結果也通過應用于體內脊髓損傷模型的細胞得以證實。
發明人發現ed-1+細胞例如活化的巨噬細胞和小神經膠質粘附于營養不良的軸突,并且這一點是回縮必須的。發明人發現將所述細胞或所述細胞的條件培養基應用于營養不良的軸突降低了或預防了粘附。發明人還發現應用培養所述細胞的條件培養基具有神經刺激作用并且顯著地增加神經突向外生長/再生。
因此,本發明主要涉及一種治療(改善或預防)與軸突回縮相關的神經元損傷的方法。
因此,本發明主要涉及一種通過促進損傷中或損傷周圍的軸突生長/再生來治療(改善或預防)神經元損傷的方法。
本發明還主要涉及一種減少神經元損傷中軸突回縮的方法。
本發明還主要涉及一種促進損傷中或損傷周圍的軸突生長/再生的方法。
回縮可由ed-1+細胞例如活化的巨噬細胞和/或小神經膠質引起。
本發明還主要涉及一種減少ed-1+細胞與營養不良的軸突的粘附從而減少軸突回縮的方法。
這些結果通過在足夠接近所述損傷的位置、以足夠的時間和足夠的量對損傷給予細胞以促進損傷中或損傷周圍的軸突生長/再生而實現。
這些結果通過在足夠接近所述損傷的位置、以足夠的時間和足夠的量對損傷給予細胞以減少神經元損傷中的軸突回縮并減少與軸突回縮相關的神經元損傷而實現。
這些結果通過在足夠接近所述損傷的位置、以足夠的時間和足夠的量對損傷給予細胞以減少ed-1+細胞與營養不良的軸突的粘附——所述粘附會導致軸突回縮——而實現。
所述細胞被引進至損傷的軸突從而所述細胞減少原有ed-1+細胞與軸突的粘附。
ed-1+細胞包括但不限于巨噬細胞和小神經膠質。
這些結果由所述細胞分泌的因子造成。因此,所述結果還通過使用條件細胞培養基或其級分或源自所述條件培養基的蛋白或其他因子而實現。所述條件培養基通過在細胞培養中培養這樣的細胞而產生,即所述細胞可有效減少粘附和軸突回縮并且/或者促進軸突生長。在一個實施方案中,所述條件培養基在用前不冷凍。
這些結果還通過使用細胞裂解物或細胞級分而實現。
可以在對應于軸突回縮及由其導致的損傷的各時間點,例如在急性損傷時給予上述細胞、分泌的因子、級分等,以在初始急性損傷后延續一段時間(例如數周)。
然而,應理解軸突回縮也可在慢性損傷情況下發生,例如下面描述的那些。在慢性損傷中,可按照任何可減少軸突回縮的方案給予所述細胞。
由于所述細胞(及分泌的因子)也會促進軸突生長,因此也可在慢性和急性損傷中給予所述細胞(及分泌的因子),所述損傷不一定與回縮相關。對這些損傷進行治療從而在損傷中或損傷周圍使得和促進軸突生長/再生。
在一個實施方案中,所述細胞為干細胞。干細胞包括但不限于胚胎干細胞和非胚胎干細胞。與胚胎干細胞一樣,非胚胎干細胞可具有分化為不止一種胚層的細胞類型和/或表達與分化為不止一種胚層的細胞類型的潛力有關的一個或多個標志物的能力。非胚胎細胞還包括組織特異性干細胞,即具有分化為僅一種胚層的細胞類型的能力的細胞,例如造血干細胞、神經干細胞和間充質干細胞。
在一個具體實施方案中,所述非胚胎干細胞已被命名為“多能成體祖細胞”("mapc")并被記載于u.s.7,015,037。
本發明涵蓋形成軸突營養不良的任何神經系統損傷,其中ed-1+細胞例如活化的巨噬細胞或小神經膠質與所述營養不良的軸突相互作用并導致所述軸突回縮。這包括中樞神經系統組織,包括腦和脊髓。與營養不良的軸突有關的病癥包括但不限于:影響脊髓的任何類型的外傷(這些包括任何來自脊髓外的力(包括椎間盤突出))或者來自脊髓內的脊髓損傷,例如脊髓空洞癥;腦內或腦外的任何類型的外傷影響產生的腦損傷(例如頭部創傷);整個中樞神經系統的中風(缺血性或溶血性的);多發性硬化;癲癇;神經變性疾病如阿耳茨海默氏病、帕金森氏病、肌萎縮側索硬化癥(葛雷克氏癥)和克羅伊茨費爾特-雅各病(cjd)。
附圖說明
圖1-脊髓損傷后再生障礙的示意圖。
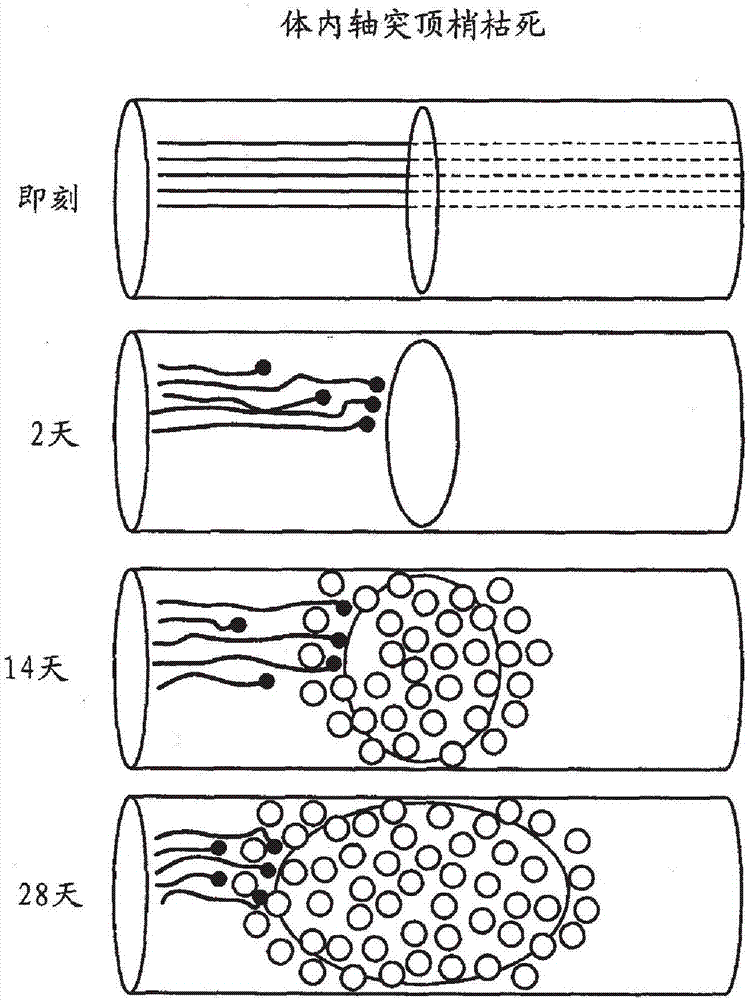
圖2-體內軸突頂梢枯死的示意圖。
圖3-背柱破碎后巨噬細胞滲入和軸突頂梢枯死的真實圖和曲線圖。巨噬細胞滲入與脊髓損傷后的軸突回縮相關。脊髓損傷后上行感覺軸突隨時間大幅回縮。所示a、b為損傷后2天(a)和7天(b)20μm厚的背柱破碎(dcc)脊髓損傷的縱向切片的組合圖。dex-tr,texasred綴合的葡聚糖3000mw。切片的方向使得尾端在圖像左側,嘴端在右側。下面的白色框表示軸突相對損傷中心(點線)的位置,同時具有來自一只動物在每個時間點的多張切片的疊加纖維描線。標尺刻度線表示200μm的增量。a,在損傷后2天,背根神經節軸突(紅色)已從損傷中心處的初始切斷位置——由有gfap+活性的星形細胞標記(藍色)——回縮了一段較短的距離。在所述損傷內存在少數ed-1+細胞(綠色),其最可能是活化的小神經膠質。b,到損傷后7天,損傷的軸突(紅色)已從損傷中心大幅回縮。現在損傷和周圍組織充滿了ed-1+細胞(綠色),其主要為滲入的巨噬細胞,而有活性的星形細胞(藍色)已離開損傷中心。c,表示平均軸突回縮隨時間的曲線圖。回縮主要發生在損傷后前7天中;然而,損傷后,回縮會一直持續,最長達28天,這是研究的時長。軸突回縮(黑色曲線圖)如下:第2天與第7、14和28天顯著不同(單因素anova,f(4,40)=6.50,p<0.001;tukey事后檢驗#p<0.05,*p<0.01,**p<0.001)。巨噬細胞清除(紅色曲線圖)如下:第2天與第7、14和28天顯著不同;第4天與第14和28天顯著不同;第7天與第2和14天顯著不同(單因素anova,f(4,40)=22.83,p<0.001;tukey事后檢驗,*p<0.01,**p<0.001,***p<0.0001)。誤差棒表示標準誤差。比例尺:a,b,250μm。
圖4-巨噬細胞體外誘導神經元頂梢枯死的縮時組合圖。在神經膠質疤痕的體外模型中,巨噬細胞誘導營養不良的成體背根神經節軸突的大幅回縮。a,縮時動畫的6幅單幀圖像的組合圖,其中將nr8383巨噬細胞加至營養不良的成體背根神經節神經元的培養物,所述神經元在生長促進胞外基質分子層粘連蛋白和強抑制性硫酸軟骨素蛋白聚糖聚集蛋白聚糖上培養。每幀的時間在每幅圖像的右下角給出,箭頭標出生長錐的中心區域。星號標記培養物中的一個固定點,作為變幀過程中的生長錐位置的參照。在30分鐘時加入巨噬細胞后巨噬細胞-生長錐接觸立即開始。在61分鐘時觀察到第二個巨噬細胞與該營養不良的軸突之間的物理接觸。當在110分鐘回縮開始時,其他巨噬細胞物理地改變了軸突軌跡。在150分鐘時,生長錐被多個巨噬細胞所遮擋并已幾乎回縮到該幀之外。比例尺,20μm。b,追蹤a中整個縮時動畫的生長錐的位置曲線圖。每一點均代表單幀(每30秒)的生長錐中心區域的位置。在接觸巨噬細胞后,軸突發生約100μm的大幅回縮。c,另一示例性縮時實驗的位置曲線圖。
圖5-軸突與巨噬細胞之間形成的接觸。在神經膠質疤痕的體外模型中,巨噬細胞與營養不良的軸突物理地相互作用。a,從巨噬細胞物理地接觸營養不良的軸突的縮時動畫中選幀。在回縮發生之前,當軸突從基底上抬起并嚴重彎曲(箭頭)時,生長錐仍然附著。b,圖3a的第三幅圖像的更高放大率的圖像。在巨噬細胞和營養不良的軸突之間發生幾處粘附性接觸。箭頭標示當巨噬細胞移開軸突時,由這些接觸形成的膜突起。c,加入巨噬細胞(綠色)后2.5小時,成體drg神經元(紅色)的培養物的40×共焦z-stack三維重構。觀察到巨噬細胞與營養不良的生長錐直接接觸。d,c圍繞x軸旋轉90°產生的所述三維重構的側視圖。箭頭標示已被臨近的巨噬細胞(綠色)從基底抬起的神經元突起(紅色)。比例尺:a,b,20μm;c,50μm。
圖6-mmp9抑制劑阻止巨噬細胞接觸所致軸突頂梢枯死的縮時組合圖。
圖7-評估外加的活細胞(mapc)或條件培養基對巨噬細胞誘導的背根神經節(drg)神經元頂梢枯死的作用的實驗設計。
圖8-與drg共培養的mapc的縮時組合圖,示出mapc的加入阻止巨噬細胞誘導的頂梢枯死。在加入巨噬細胞前的1天給予mapc。
圖9-實驗縮時組合圖,顯示mapc條件培養基阻止巨噬細胞誘導的軸突頂梢枯死。在加入條件培養基30分鐘后加入巨噬細胞。
圖10-顯示以mapc條件培養基刺激的巨噬細胞不誘導軸突頂梢枯死的縮時組合圖。
圖11-mapc阻止巨噬細胞介導的軸突頂梢枯死的圖示。
圖12-用mapc或條件培養基的體外實驗的實驗總結。
圖13-mapc阻止背柱破碎損傷后巨噬細胞介導的軸突頂梢枯死并促進再生進入損傷中心。移植了媒介(vehicle)對照或mapc的損傷后7天脊髓切片的曲線圖和真實圖。在神經膠質疤痕的體外模型中,巨噬細胞誘導營養不良的成體背根神經節軸突的大幅回縮。將nr8383巨噬細胞加至營養不良的成體背根神經節神經元的培養物,所述神經元在生長促進胞外基質分子層粘連蛋白和強抑制性硫酸軟骨素蛋白聚糖聚集蛋白聚糖的相反斑點梯度上培養。位置圖追蹤整個縮時動畫的生長錐。每一點代表單幀(每30秒)的生長錐中心區域的位置。在接觸巨噬細胞后,軸突發生約100μm的大幅回縮。
畫面示出了損傷后7天20μm厚的背柱破碎(dcc)脊髓損傷的縱向切片的10×組合圖。以texasred綴合的3000mw葡聚糖標記纖維,并且以ed-1+(紫色)可視化巨噬細胞。切片的方向使得尾端在圖像左側,嘴端在右側。損傷中心標記在下面(黑實線),同時具有來自一只動物在每個時間點的多張切片的3條疊加纖維描線。a,在損傷后7天并且在只注射媒介的情況下,背根神經節軸突(紅色)已從損傷中心處的初始切斷位置大幅回縮了一段距離。b,到損傷后7天并且在mapc移植物的情況下,損傷的軸突(紅色)已大量再生進入損傷。c,表示在接受媒介對照或mapc移植物的動物在損傷后2、4和7天的平均軸突回縮的圖表。通過一般線性模型,所述條件、mapc移植物與媒介對照顯著不同,*p<0.0001。比例尺:a,b,200μm。
圖14-顯示ng2+神經膠質細胞不阻止巨噬細胞誘導的頂梢回縮的縮時組合圖。ng2+細胞可使軸突穩定,但在體外不阻止巨噬細胞攻擊后巨噬細胞介導的回縮。a,顯示體外2日后在聚集蛋白聚糖和層粘連蛋白的梯度上β微管蛋白+(紅色)軸突與ng2+(綠色)細胞的軸突聯系的40×共焦圖像。邊緣以白色點線標示。b,ng2+細胞表達波形蛋白(紅色)。c,ng2+細胞表達巢蛋白(nestin)(紅色)。d,顯示在成體小鼠脊髓ng2+細胞的存在下在聚集蛋白聚糖/層粘連蛋白的梯度上巨噬細胞/軸突的相互作用的縮時動畫中的6幀示例性圖像。將nr8383巨噬細胞加至成體drg神經元的2div培養物中。每幀的時間在每幅圖像的右下角給出,星號標記培養皿上的一個固定點,作為變幀過程中位置的參照。箭頭標示生長錐的中心區域。觀察30分鐘后加入巨噬細胞,第一次接觸發生在103分鐘。到110分鐘,軸突已經發生長距離的回縮。空心箭頭標示回縮纖維的存在。e,d中所示的延縮時動畫的每幀(30秒)的生長錐位置的曲線圖。紅色弧線代表所述斑點的內邊緣的位置。箭頭表示初始生長軌跡。f,在集聚蛋白聚糖/層粘連蛋白的斑點梯度上與ng2+細胞的共培養的6個營養不良的軸突在與巨噬細胞接觸后距離原點的距離。箭頭標示軸突已經回縮至ng2細胞的位置。比例尺:a,b,c,50μm。d,20μm。
圖15-在斑點梯度上單獨培養的mapc的共焦圖像以及在斑點梯度上與神經元一起培養的mapc的更高放大率的圖像。
在集聚蛋白聚糖——通過cs56(紅色)可視化——和層粘連蛋白的雙向梯度上培養的gfp+mapc(綠色)的10×共焦圖像。與通過β微管蛋白(藍色)可視化的成體drg神經元共培養的mapc的40×共焦圖像。在體外2日后,成體drg和mapc均不通過抑制性斑點邊緣。
加至集聚蛋白聚糖斑點梯度的mapc未進入抑制性邊緣,但良好地粘附于斑點中心并與成體drg軸突連接。
圖16-對照培養基或mapc條件培養基在體外對軸突向外生長的作用的曲線圖和真實圖。
圖17-顯示對照培養基不阻止巨噬細胞誘導的頂梢枯死的實驗的縮時組合圖。在加入條件培養基30分鐘后加入巨噬細胞。
雖然存在對照mapc培養基,但是在神經膠質疤痕的體外模型中,巨噬細胞仍誘導營養不良的成體背根神經節軸突的大幅回縮。a,縮時動畫的6幅單幀圖像的組合圖,其中將nr8383巨噬細胞加至營養不良的成體背根神經節神經元的培養物,所述神經元在生長促進胞外基質分子層粘連蛋白和強抑制性硫酸軟骨素蛋白聚糖聚集蛋白聚糖的相反斑點梯度上培養。每幀的時間在每幅圖像的右下角給出,箭頭標出生長錐的中心區域。星號標記培養物中的一個固定點,作為變幀過程中的生長錐位置的參照。比例尺,20μm。b,追蹤a中整個縮時動畫的生長錐的位置曲線圖。每一點代表單幀(每30秒)的生長錐中心區域的位置。在接觸巨噬細胞后,軸突發生約80μm的大幅回縮。
直接向所述縮時培養皿中加入mapc條件培養基導致了生長錐形態的改變,從營養不良的、停滯的狀態變為能動的、扁平的狀態。巨噬細胞仍然接觸這些軸突,但接觸通常是瞬時的并且通常不導致軸突回縮。對照mapc培養基不阻止軸突回縮。以mapc條件培養基預處理的巨噬細胞也在所述斑點上接觸軸突,但不引起回縮(圖9-12)。mapc可能作用于巨噬細胞,從而改變它們的受體表達、對損傷細胞的響應或者mmp-9的分泌。
具體實施方式
定義
"a"或"an"在本文中是指一個或多個;至少一個。當本文中使用復數形式時,通常包括單數形式。
本文使用的術語“粘附、附著、粘著”等,是指連接足夠長的時間以誘導軸突回縮。如本文進一步描述的,應理解物理接觸可發生在巨噬細胞(或其他細胞)與營養不良的軸突之間,所述接觸是短暫的并且不導致軸突回縮。在本發明的上下文中,被本發明試劑降低或阻止的附著是持續足夠長的時間以誘導軸突回縮的附著。因此,本發明不排除允許營養不良的軸突與ed-1+細胞之間物理接觸的試劑。因此,本發明涵蓋允許接觸(例如短暫的物理接觸)但不允許附著足夠長的時間以導致軸突頂梢枯死的試劑。
“共給予”是指彼此結合、一起、配合地給予,包括同時或依次給予兩種或多種試劑。
在沒有其他限制的情況下,“包含”是指必須包括所指事物,但不對其他可包括的事物進行限制或排除。例如,“包含x和y的組合物”包括含有x和y的任何組合物,而不管所述組合物中是否存在其他組分。同樣,“包含步驟x的方法”包括其中進行x(不管x是所述方法中的唯一步驟還是只是步驟之一)的任何方法,不管可能存在多少其他步驟,也不管x與這些步驟相比多么簡單或復雜。“包括”和使用詞根“含”的類似短語在本文中用作“包含”的同意詞并具有相同的含義。
“包括”是“包含”的同義詞(見上)。
“條件細胞培養基”是本領域公知的術語,指已經培養過細胞的培養基。在本文中,這是指將所述細胞培養足夠的時間以分泌對減少活化的巨噬細胞與營養不良的神經元的粘附和/或促進神經突向外生長/軸突再生有效的因子。
條件細胞培養基是指細胞已經在其中培養從而將因子分泌到所述培養基中的培養基。出于本發明的目的,可使細胞生長歷經足夠數量的細胞分裂以產生有效量的所述因子,使得所述培養基減少巨噬細胞與營養不良的神經元的粘附并因此減少軸突回縮和/或促進神經突向外生長/軸突再生。通過本領域任何已知的方法將細胞從所述培養基中除去,所述方法包括但不限于離心、過濾、免疫清除(例如通過標記的抗體和磁性柱)和facs分選。
“頂梢枯死”是用于指因軸突損傷而發生的軸突回縮的技術術語。在本發明上下文中,軸突回縮是指因ed-1+細胞并且特別是巨噬細胞和小神經膠質的充分粘附而發生的回縮。所述巨噬細胞和小神經膠質(即ed-1+細胞)不處于休眠或不活動狀態。它們是被活化的。術語“活化的”是指這些細胞允許其粘附于營養不良的軸突從而導致軸突回縮的狀態。導致體外活化的條件的實例在本申請中有進一步描述。然而,應理解所述活化不限于本文公開的具體條件。
盡管本發明通常具體涉及活化的巨噬細胞(并以其舉例),然而這些細胞是一類ed-1+細胞并且本發明也適用于其他這類細胞。一個實例是活化的小神經膠質。
“有效量”通常指提供需要的局部或全身作用的量。例如,有效量是足以實現有益或需要的臨床效果的量。所述有效量可以單次給藥的形式一次全部提供或以多次給藥的形式分份提供。對什么量會被認為是有效量的準確確定可基于每個受試者的個人因素,包括他們的身高體重、年齡、損傷和/或待治療的疾病或損傷以及遭受損傷或疾病開始后的時間。本領域技術人員能夠基于這些本領域中慣常的考慮確定給定受試者的有效量。當用于本文時,“有效劑量”與“有效量”所指相同。
“有效途徑”通常是指提供用于將一種試劑送遞至所需腔室、系統或位置的途徑。例如,有效途徑是給予試劑以在所需的作用位點提供量足以實現有益或所需的臨床效果的所述試劑的途徑。
“ec細胞”是從被稱為畸胎瘤的一種癌癥的分析中發現的。1964年,研究人員注意到畸胎瘤中的單個細胞可被分離并在培養中保持不分化狀態。于是這種類型的干細胞被稱為胚胎癌細胞(ec細胞,embryoniccarcinomacell)。
“胚胎干細胞(esc)”為本領域所熟知并多年以來已從多種不同哺乳動物種中制得。胚胎干細胞是源自被稱為胚泡的早期胚胎的內細胞團的干細胞。它們能夠分化為三種原胚層的所有衍生細胞:外胚層、內胚層和中胚層。這些包括成體內超過220種細胞類型的每一種。es細胞可成為體內除胎盤外的任何組織。只有桑椹胚細胞是全能性的,能夠成為所有組織和胎盤。
使用術語“包括”并非意圖進行限制。例如,述及抗體抑制劑“包括”片段和變體時不表示排除其他形式的抗體抑制劑。
“誘導的多能干細胞(ipsc或ips細胞)”是例如通過引入外源基因被再編程的體細胞,其中所述外源基因賦予所述體細胞分化程度較低的表型。然后這些細胞可被誘導分化為分化程度較低的后代。使用首先在2006年公開的方法的改進方法獲得了ips細胞(yamanaka,s.etal.,cellstemcell,1:39-49(2007))。例如,在一個實例中,為獲得ips細胞,科學家以皮膚細胞為起始材料,通過用反轉錄病毒將基因插入到細胞dna中的標準實驗室技術隨后對所述皮膚細胞進行了改造。在一個實例中,所插入的基因為oct4、sox2、lif4和c-myc,已知這些基因作為天然調控因子一起作用以使細胞保持在與胚胎干細胞相似的狀態。文獻已對這些細胞進行了描述。參見,例如,wernigetal.,pnas,105:5856-5861(2008);jaenischetal.,cell,132:567-582(2008);hannaetal.,cell,133:250-264(2008);以及brambrinketal.,cellstemcell,2:151-159(2008)。這些參考文獻以引用的方式納入以教導ipsc和制備它們的方法。也可通過特定培養條件(暴露于特定試劑)獲得這些細胞。
術語“分離的”指不與一種或多種細胞或者在體內時與所述一種細胞或多種細胞結合的一種或多種細胞組分結合的一種或多種細胞。“富集群落”指合乎需要的細胞相對體內或原代培養物中的一種或多種其他細胞類型的數量上相對增加。
然而,本文所用的術語“分離的”不表示只存在胚胎干細胞。而是,術語“分離的”表示細胞從其天然組織環境中取出并以相對所述正常組織環境更高的濃度存在。因此,“分離的”細胞群落可進一步包括干細胞以外的細胞類型并可包括其他組織組分。這可用例如細胞的倍增來表示。細胞可在體外或離體進行10、20、30、40次或更多次的倍增,從而相對其在體內或在原始組織環境(例如骨髓、外周血、脂肪組織等)中的原始數量被富集。
“mapc”是“多能成體祖細胞”的首字母縮寫。其指非胚胎干細胞。mapc中的術語“成體”是非限制性的。其指非胚胎體細胞。與胚胎干細胞一樣,mapc可產生多于一種胚層的細胞譜系。其可在分化時產生全部三種胚層(即內胚層、中胚層和外胚層)的細胞類型。與胚胎干細胞類似,人mapc表達端粒末端轉移酶、oct3/4(即oct3a)、rex-1、rox-1和sox-2,并可表達ssea-4。(亦見jiang,y.etal.,nature,418:41(2002);exphematol,30:896(2002))。端粒在mapc中延長并且核型正常。由于注射到哺乳動物中的mapc可遷移至多個器官內并在這些器官內被同化,因此mapc是自我更新的干細胞。對于mapc,“多能”指在分化時產生多于一種原始胚層(即內胚層、中胚層和外胚層)例如全部三種胚層的細胞系的能力。
“神經突向外生長”是指損傷位置處的神經元不僅停止回縮而且生長并延長的性質。
“可藥用載體”是本發明中所用細胞的任何可藥用介質。所述介質可保持等滲性、細胞代謝、ph等。所述介質與對受試者的體內給藥相容,并因此可用于細胞遞送和治療。
可培養并刺激“原始胚胎生殖細胞”(pg或eg細胞)以產生許多分化程度較低的細胞類型。
“祖細胞”是在干細胞分化過程中產生的具有其最終分化后代的某些但非全部特征的細胞。限定的祖細胞——例如“心肌祖細胞”——定向到細胞系,但不定向到特定的或最終分化的細胞類型。當用于縮寫"mapc"時,術語“祖”不將這些細胞限定到具體的細胞系。
本文使用的術語“減少”是指阻止以及減輕。當用于治療的語境時,“減少”是阻止或改善一種或多種臨床癥狀。臨床癥狀是一種(或多種)對或將對——如果不進行治療——受試者的生命質量(健康)有負面影響的癥狀。
術語“回縮”是指軸突逐漸遠離損傷位置例如神經膠質疤痕形成之處。此處,再生軸突的末端停止延長并出現營養不良。然后,這些營養不良的末端可進一步逐漸遠離神經膠質疤痕和損傷位置。
“自我更新”指復制出具有與親代細胞相同的分化潛能的子代干細胞的能力。在本文中使用的相似的術語是“增殖”。
“干細胞”指可進行自我更新(即后代具有相同的分化潛能)并且也可產生分化潛能受到更多限制的后代細胞的細胞。在本發明的上下文中,干細胞還可包括已經例如通過核移植、通過與更原始的干細胞融合、通過引入特定轉錄因子或通過在特定條件下培養而已去分化的分化程度更高的細胞。參見,例如wilmutetal.,nature,385:810-813(1997);yingetal.,nature,416:545-548(2002);guanetal.,nature,440:1199-1203(2006);takahashietal.,cell,126:663-676(2006);okitaetal.,nature,448:313-317(2007);以及takahashietal.,cell,131:861-872(2007)。
去分化也可通過給予某些化合物或暴露于可導致去分化的體外或體內物理環境下而導致。干細胞還可源自異常組織,例如畸胎瘤,以及某些其他來源,例如擬胚體(雖然擬胚體可被認為是胚胎干細胞,因為它們源自胚胎組織,盡管不直接來自內細胞團)。干細胞還可通過將與干細胞功能相關的基因引入非干細胞中而產生,例如誘導的多能干細胞。
“受試者”是指脊椎動物,例如哺乳動物,例如人。哺乳動物包括但不限于人、狗、貓、馬、牛和豬。
術語“治療有效量”指被確定在哺乳動物中產生任何治療性反應的量。例如,有效量的治療性細胞或細胞相關劑可延長患者的存活和/或抑制明顯的臨床癥狀。具有治療效果的治療,在該術語用于本文時的含義下,包括改善受試者生命質量的治療,即使所述治療不會改善疾病本身的后果。這種治療有效量由本領域普通技術人員容易地確定。因此,“治療”是指送遞所述量。因此,治療可阻止或緩解因活化的巨噬細胞與營養不良的軸突的粘附而發生的任何病理癥狀。治療還指軸突再生的有益臨床效果。
“治療”在本發明中廣泛地使用,這一術語包括預防、緩解、抑制或治愈缺陷、功能障礙、疾病或其他有害過程等,包括干擾治療和/或由治療引起的過程。
干細胞
可實施本發明,優選地使用脊椎動物物種例如人、非人靈長類、家畜、牲畜和其他非人哺乳動物的干細胞實施本發明。這些細胞包括但不限于下文所述的細胞。
胚胎干細胞
研究最深入的干細胞是胚胎干細胞(esc),因為其具有無限自我更新和多能分化潛能。這些細胞源自胚泡的內細胞團或者可源自著床后胚胎的原始生殖細胞(胚胎生殖細胞或eg細胞)。es和eg細胞已首先從小鼠中得到,后來從許多不同動物得到,最近還從非人靈長類和人中得到。當引入小鼠胚泡或其他動物的胚泡中時,esc可促使所述動物的所有組織的形成。es和eg細胞可通過用抗ssea1(小鼠)和ssea4(人)的抗體進行陽性染色而被識別。參見例如美國專利no.5,453,357;5,656,479;5,670,372;5,843,780;5,874,301;5,914,268;6,110,7396,190,910;6,200,806;6,432,711;6,436,701,6,500,668;6,703,279;6,875,607;7,029,913;7,112,437;7,145,057;7,153,684和7,294,508,所述專利的每一篇都以引用的方式納入本文以教導胚胎干細胞及其制備和擴增方法。因此,esc及其分離和擴增方法為本領域所熟知。
若干影響體內胚胎干細胞潛力狀態的轉錄因子和外源細胞因子已被鑒定出。將要描述的與干細胞多能性相關的第一種轉錄因子是oct4。oct4屬于pou(pit-oct-unc)家族的轉錄因子并且是能夠活化在啟動子或增強子區內的含有被稱為“八聚體基序”的八聚體序列的基因轉錄的dna結合蛋白。oct4在受精卵的卵裂期表達直至卵圓筒形成。oct3/4的功能是抑制誘導分化的基因(即foxad3、hcg)以及活化促進多能性的基因(fgf4、utf1、rex1)。sox2——高遷移率組(hmg)盒轉錄因子的一員——與oct4共同作用以活化在內細胞團中表達的基因的轉錄。將oct3/4在胚胎干細胞中的表達維持某些水平之間是必需的。oct4表達水平>50%的過表達或下調會改變胚胎干細胞的命運,分別形成原始內胚層/中胚層或滋養外胚層。在體內,oct4缺陷胚胎發育至胚泡期,但內細胞團的細胞不具有多能性。相反,它們沿胚外滋養層譜系分化。sall4——一種哺乳動物spalt轉錄因子——是oct4的上游調節子,從而對在胚胎發生早期維持oct4的適當水平起重要作用。當sall4下降到低于某個閾值時,滋養外胚層細胞會錯位擴張到內細胞團中。多能性所需的另一轉錄因子是nanog,以一個凱爾特人部落"tirnanog":永遠年輕的土地命名。在體內,nanog從致密桑椹胚期表達,隨后被限定于內細胞團并在著床期下調。nanog的下調可對于防止多能性細胞不受控制的擴增以及允許原腸胚形成期間的多譜系分化起重要作用。在5.5天分離的無nanog的胚胎由無組織的胚泡構成,主要包含胚外內胚層,而不含可辨別的上胚層。
非胚胎干細胞
干細胞已在大多數組織中被發現。也許表征最好的是造血干細胞(hsc)。hsc是源自中胚層的細胞,其可使用細胞表面標志物和功能性特征進行純化。它們已從骨髓、外周血、臍帶血、胎肝和卵黃囊中分離。它們發起血細胞生成并產生多個造血譜系。當移植到受到致死輻射的動物中時,它們可重建紅血球、中性白細胞-巨噬細胞、巨核細胞和淋巴造血細胞池。它們也可被誘導已進行某些自我更新細胞分裂。參見例如美國專利no.5,635,387;5,460,964;5,677,136;5,750,397;5,681,599和5,716,827。美國專利no.5,192,553報道了分離人新生兒或胎兒造血干細胞或祖細胞的方法。美國專利no.5,716,827報道了為thy-1+祖細胞的人造血細胞和在體外再生所述細胞的合適的生長培養基。美國專利no.5,635,387報道了培養人造血細胞及其前體的方法和設備。美國專利no.6,015,554記載了重構成人淋巴和樹突細胞的方法。因此,hsc及其分離和擴增方法為本領域所熟知。
本領域熟知的另一種干細胞是神經干細胞(nsc)。這些細胞可在體內增殖并持續再生至少某些神經細胞。當離體培養時,神經干細胞可被誘導進行增殖以及分化為不同類型的神經元和神經膠質細胞。當移植到腦中時,神經干細胞可定植并產生神經細胞和神經膠質細胞。參見,例如gagef.h.,science,287:1433-1438(2000),svendsens.n.etal,brainpathology,9:499-513(1999),和okabes.etal.,mechdevelopment,59:89-102(1996)。美國專利no.5,851,832報道了從腦組織獲取的多能性神經干細胞。美國專利no.5,766,948報道了從新生大腦半球產生成神經細胞。美國專利no.5,564,183和5,849,553報道了哺乳動物神經嵴干細胞的用途。美國專利no.6,040,180報道了從哺乳動物多能性cns干細胞培養物體外產生分化的神經元。wo98/50526和wo99/01159報道了神經上皮干細胞、少突膠質細胞-星形細胞前體和譜系限制性神經元前體的產生和分離。美國專利no.5,968,829報道了從胚胎前腦獲取的神經干細胞。因此,神經干細胞及其制備和擴增方法為本領域所熟知。
本領域廣泛研究的另一種干細胞是間充質干細胞(msc)。msc來源于胚胎中胚層并可從許多來源分離,包括成體骨髓、外周血、脂肪、胎盤、臍帶血等。msc可分化為許多中胚層組織,包括肌肉、骨、軟骨、脂肪和腱。有大量關于這些細胞的文獻。參見例如美國專利no.5,486,389;5,827,735;5,811,094;5,736,396;5,837,539;5,837,670;和5,827,740。也參見pittenger,m.etal,science,284:143-147(1999)
成體干細胞的另一實例是脂肪來源的成體干細胞(adsc),其已從脂肪分離,通常是通過吸脂術繼而使用膠原酶釋放adsc。adsc在許多方面類似于來源于骨髓的msc,除了可從脂肪分離更多的細胞之外。這些細胞已被報告可分化為骨、脂肪、肌肉、軟骨和神經元。一種分離方法記載于u.s.2005/0153442。
本領域已知的其他干細胞包括胃腸干細胞、表皮干細胞和肝干細胞,其中肝干細胞也被稱為“卵圓細胞”(potten,c,etal.,transrsoclondbbiolsci,353:821-830(1998),watt,f.,transrsoclondbbiolsci,353:831(1997);alisonetal.,hepatology,29:678-683(1998))。
被報道能夠分化為一種以上胚層細胞類型的其他非胚胎細胞包括但不限于來自臍帶血(參見美國專利公開no.2002/0164794)、胎盤(參見美國專利公開no.2003/0181269)、臍帶基質(mitchelletal.,stemcells,21:50-60,2003)、小胚胎樣干細胞(kuciaetal.,jphysiolpharmacol,57suppl5:5-18,2006)、羊水干細胞(atala,a.,jtissueregenmed1:83-96,2007)、源自皮膚的前體細胞(tomaetal.,natcellbiol3:778-784,2001)以及骨髓(參見美國專利公開no.2003/0059414和2006/0147246)的細胞,上述文獻的每一篇都以引用的方式納入本文中以教導這些細胞。
對體細胞再編程的方法
若干不同方法,例如核移植、細胞融合和培養誘導的再編程,已用于誘導已分化細胞轉化為胚胎狀態。核移植包括將體細胞核注射到去核卵母細胞中,若將所述卵母細胞轉移到代孕母體中,其可長成克隆(“生殖性克隆”),或者,若外植培養可長成遺傳匹配的胚胎干(es)細胞("體細胞核移植",scnt)。體細胞與es細胞的細胞融合產生具有多能es細胞全部特征的雜交細胞。體細胞的外植培養選擇出可具有多能性的永生細胞系。目前,精原干細胞是可從出生后動物中得到的多能細胞的唯一來源。用限定的因子轉導體細胞可啟動產生多能狀態的再編程。這些實驗方法已被廣泛地綜述(hochedlingerandjaenisch,nature441:1061-1067(2006)和yamanaka,s.,cellstemcell1:39-49(2007))。
核移植
核移植(nt),也稱為體細胞核移植(scnt),是指將供體體細胞的細胞核引入去核卵母細胞中以長成克隆動物例如多莉羊(wilmutetal.,nature385:810-813(1997))。通過nt實現的活體動物的產生表明了體細胞(包括最終分化的細胞)的外遺傳狀態是穩定的,但并非被不可逆地固定,而是可被再編程為胚胎狀態,所述胚胎狀態能夠指導新生物體的發育。除了提供闡明胚胎發育和疾病涉及的基本外遺傳機制的令人激動的實驗方法之外,核克隆技術還可能引起患者特異性移植醫學的興趣。
體細胞和胚胎干細胞的融合
體細胞核到未分化狀態的外遺傳再編程已在通過胚胎干細胞與體細胞的融合產生的鼠雜種中被證明。多種體細胞和胚胎癌細胞、(solter,d.,natrevgenet,7:319-327(2006)、胚胎生殖細胞(eg)或es細胞(zwakaandthomson,development,132:227-233(2005))的雜交細胞與親代胚胎細胞具有許多相同的性質,表明在這些融合產物中多能性表型是顯性的。與小鼠(tadaetal.,currbiol,11:1553-1558(2001))一樣,人es細胞具有在融合后對體細胞核再編程的潛力(cowanetal.,science,309:1369-1373(2005));yuetal.,science,318:1917-1920(2006))。沉默多能性標志物例如oct4的活化或失活體x染色體的再活化為雜交細胞中體基因組的再編程提供了分子證據。已經表明dna復制對于多能性標志物——其在融合后2天首次觀察到——的活化是必需的(doandscholer,stemcells,22:941-949(2004));并且當與神經干細胞融合時,es細胞中nanog的強制過表達促進多能性(silvaetal.,nature,441:997-1001(2006))。
培養誘導的再編程
已從胚胎來源獲得了多能細胞,所述胚胎來源例如胚泡的卵裂球和內細胞團(icm)(es細胞)、上胚層(episc細胞)、原始生殖細胞(eg細胞)和出生后精原干細胞("magscsm"“es樣”細胞)。多能細胞以及它們的供體細胞/組織如下:單性生殖es細胞源自鼠卵母細胞(narasimhaetal.,currbiol,7:881-884(1997));胚胎干細胞源自卵裂球(wakayamaetal.,stemcells,25:986-993(2007));內細胞團細胞(來源不適用)(egganetal.,nature,428:44-49(2004));胚胎生殖細胞和胚胎癌細胞源自原始生殖細胞(matsuietal.,cell,70:841-847(1992));gmcs、massc和masc源自精原干細胞(guanetal.,nature,440:1199-1203(2006);kanatsu-shinoharaetal.,cell,119:1001-1012(2004);和seandeletal.,nature,449:346-350(2007));episc細胞源自上胚層(bronsetal.,nature,448:191-195(2007);tesaretal.,nature,448:196-199(2007));單性生殖es細胞源自人卵母細胞(cibellietal.,science,295l819(2002);revazovaetal.,cloningstemcells,9:432-449(2007));人es細胞源自人胚泡(thomsonetal.,science,282:1145-1147(1998));mapc源自骨髓(jiangetal.,nature,418:41-49(2002);phinneyandprockop,stemcells,25:2896-2902(2007));臍帶血細胞(源自臍帶血)(vandevenetal.,exphematol,35:1753-1765(2007));神經球源細胞源自神經細胞(clarkeetal.,science,288:1660-1663(2000))。已知來自生殖細胞系的供體細胞例如pgc或精原干細胞在體內是單能性的,然而已證明可在延長體外培養后分離多能es樣細胞(kanatsu-shinoharaetal.,cell,119:1001-1012(2004))或magsc(guanetal.,nature,440:1199-1203(2006))。盡管這些多能細胞類型中的大多數能夠在體外分化并形成畸胎瘤,然而按照更嚴格的標準,只有es、eg、ec和源自精原干細胞的magcs或es樣細胞是多能性的,因為它們能夠形成出生后嵌合體并形成種系。最近,從成體小鼠的睪丸精原干細胞獲得了多能成體精原干細胞(masc),這些細胞的表達譜不同于es細胞(seandeletal.,nature,449:346-350(2007)),但類似于episc細胞,episc細胞源自著床后小鼠胚胎的上胚層(bronsetal.,nature448:191-195(2007);tesaretal.,nature,448:196-199(2007))。
通過限定的轉錄因子進行的再編程
takahashi和yamanaka已經報道了將體細胞再編程回es樣狀態(takahashiandyamanaka,cell,126:663-676(2006))。在四種轉錄因子oct4、sox2、c-myc和klf4的病毒介導的轉導繼而就oct4靶向基因fbxl5的活化進行篩選后,他們成功地將小鼠胚胎成纖維細胞(mef)和成體成纖維細胞再編程為多能es樣細胞(圖2a)。已活化fbxl5的細胞被稱為ips(誘導的多能干)細胞并通過其形成畸胎瘤的能力顯示具有多能性,盡管它們不能產生活嵌合體。此多能性狀態倚賴轉導的oct4和sox2基因的持續病毒表達,而內源oct4和nanog基因不表達或以較es細胞中為低的水平表達,并且它們各自的啟動子被發現在很大程度上甲基化。這與fbxl5-ips細胞與es細胞不相應但可表現不完全再編程狀態的結論一致。盡管遺傳實驗已證明oct4和sox2對于多能性是必需的(chambersandsmith,oncogene,23:7150-7160(2004);ivanonaetal.,nature,442:5330538(2006);masuietal.,natcellbiol,9:625-635(2007)),然而兩種致癌基因c-myc和klf4在再編程中的作用尚不清楚。實際上,這些致癌基因中的某些可能對于再編程是非必需的,因為小鼠和人ips細胞均已在不進行c-myc轉導的情況下得到,盡管效率較低(nakagawaetal.,natbiotechnol,26:191-106(2008);werningetal.,nature,448:318-324(2008);yuetal.,science,318:1917-1920(2007))。
mapc
mapc是“多能成體祖細胞”(非es、非eg、非生殖細胞)的首字母縮寫。mapc具有分化為至少兩種例如全部三種原始胚層(外胚層、中胚層和內胚層)的細胞類型的能力。在mapc中也發現了在es細胞中發現的基因(例如端粒末端轉移酶、oct3/4、rex-1、rox-1、sox-2)。oct3/4(在人中為oct3a)似乎是es和生殖細胞特異的。mapc代表一種比msc更原始的祖細胞群體,并且具有包括上皮、內皮、神經、肌原、生血、生骨、肝原、軟骨形成和生脂的細胞系的分化能力(verfaillie,cm.,trendscellbiol12:502-8(2002),jahagirdar,b.n.,etal.,exphematol,29:543-56(2001);reyes,m.andcm.verfaillie,annnyacadsci,938:231-233(2001);jiang,y.etal.,exphematol,30896-904(2002);和jiang,y.etal.,nature,418:41-9.(2002))。
美國專利7,015,037和美國專利申請no.10/467,963記載了人mapc。在其他哺乳動物中已鑒定出mapc。例如,美國專利7,015,037和美國專利申請no.10/467,963還記載了鼠mapc。美國專利申請no.10/467,963還記載了大鼠mapc。
這些文獻記載由catherineverfaillie分離的mapc的內容以引用的方式納入本文。
mapc的分離和生長
mapc分離方法為本領域所知。參見,例如美國專利7,015,037和美國專利申請no.10/467,963,這些方法連同mapc的表征(表型)以引用的方式納入本文。mapc可從多種來源中分離,所述來源包括但不限于骨髓、胎盤、臍帶和臍帶血、肌肉、腦、肝臟、脊髓、血液或皮膚。因此,可得到骨髓抽出物、腦或肝活組織以及其他器官,并使用本領域技術人員可采用的陽性或陰性選擇技術、根據在這些細胞中表達(或不表達)的基因分離所述細胞(例如通過功能或形態測定例如上面引用的申請中公開的那些,所述申請以引用的方式納入本文)。
如美國專利7,015,037所述的來自人骨髓的mapc
mapc不表達共同白細胞抗原cd45或成紅細胞特異性血型糖蛋白-a(gly-a)。對混合的細胞群體進行聚蔗糖-泛影葡胺分離。然后使用抗cd45和抗gly-a的抗體對所述細胞進行陰性選擇,清除cd45+和gly-a+細胞群體,然后回收剩余的大約0.1%的骨髓單核細胞。也可將細胞鋪板于纖連蛋白包被的孔中并如下所述培養2-4周以清除cd45+細胞和gly-a+細胞。在貼壁骨髓細胞的培養中,許多貼壁基質細胞在細胞倍增約30次發生復制衰老,更純系的細胞群體繼續擴增并保持長端粒。
或者,陽性選擇可用于通過細胞特異性標志物的結合來分離細胞。陽性和陰性選擇技術對本領域技術人員都是可用的,適于陰性選擇目的的多種單克隆和多克隆抗體也是本領域可獲得的(參見,例如leukocytetypingv,schlossman,etal.,eds.(1995)oxforduniversitypress)并可從若干來源購得。
從細胞群體的混合物中分離哺乳動物細胞的技術還在schwartz等人的美國專利no.5,759,793(磁性分離)、baschetal.,1983(免疫親和色譜)以及wysockiandsato,1978(熒光活化的細胞分選)中有所記載。
如美國專利7,015,037所述培養mapc
如本文所述分離的mapc可使用本文和美國專利7,015,037公開的方法培養,該專利以引用的方式納入本文以教導這些方法。
其他培養方法
在另外的實驗中,mapc的培養密度可從約100個細胞/cm2到約150個細胞/cm2到約10,000個細胞/cm2變化,包括約200個細胞/cm2到約1500個細胞/cm2到約2000個細胞/cm2。所述密度可因物種不同而變化。此外,最佳密度可根據培養條件和細胞來源而變化。為給定的培養條件和細胞確定最佳密度在本領域普通技術人員的能力范圍之內。
此外,在培養中mapc的分離、生長和分化過程中的任意時間都可使用低于約10%,包括約1-5%,特別是3-5%的有效大氣氧濃度。
細胞可在不同血清濃度下培養,例如約2-20%。可使用胎牛血清。較高的血清——例如約15-20%——可結合較低的氧壓使用。在附著于培養皿之前不需選擇細胞。例如,在聚蔗糖梯度離心后,可將細胞直接鋪板,例如250,000-500,000/cm2。可挑取、可能合并并且擴增附著的克隆。
在實施例的實驗過程中使用的一個實施方案中,使用了高血清(約15-20%)和低氧(約3-5%)的條件進行細胞培養。具體地,將來自集落的附著細胞在18%血清和3%氧氣(含有pdgf和egf)中以約1700-2300細胞/cm2的密度鋪板并傳代。
在針對mapc的實施方案中,添加物為可使mapc保留分化為全部三種細胞譜系的能力的細胞因子或組分。這可通過未分化狀態的特異性標志物的表達來指示。例如,mapc組成型地表達oct3/4(oct3a)并保持高水平的端粒末端轉移酶。
細胞培養
通常,用于本發明的細胞可在本領域可獲得并熟知的培養基中維持和擴增。這些培養基包括但不限于dulbecco'smodifiedeagle's
也考慮用哺乳動物血清補充細胞培養基。血清通常含有對存活和擴增必需的細胞因子和組分。血清的實例包括胎牛血清(fbs)、牛血清(bs)、小牛血清(cs)、小胎牛血清(fcs)、新生小牛血清(ncs)、山羊血清(gs)、馬血清(hs)、人血清、雞血清、豬血清、綿羊血清、兔血清、血清替代物和牛胚胎液(embryonicfluid)。應理解,如果認為必須滅活補體級聯的組分,那么可在55-65℃加熱滅活血清。
也可有利地使用其他補充物來為細胞提供必需的微量元素以優化生長和擴增。這些補充物包括胰島素、轉鐵蛋白、硒化鈉及其結合物。這些組分可包含在鹽溶液中,例如但不限hanks'balancedsalt
激素也可有利地用于細胞培養,包括但不限于d-醛甾酮、乙酚(des)、地塞米松、β-雌二醇、氫化可的松、胰島素、催乳素、孕酮、生長抑素/人生長激素(hgh)、促甲狀腺素、甲狀腺素和l-甲狀腺素。
根據細胞類型和分化細胞的命運,還可使用脂質和脂質載體來補充細胞培養基。這類脂質和載體可包括但不限于環糊精(α、β、γ)、膽固醇、與白蛋白綴合的亞油酸、與白蛋白綴合的亞油酸和油酸、非綴合的亞油酸、與白蛋白綴合的亞油酸-油酸-花生四烯酸、非綴合的與白蛋白綴合的油酸等。
也考慮使用飼養細胞層。飼養細胞用于支持需要復雜營養的培養細胞特別是es細胞的生長。飼養細胞是通過γ輻射滅活的正常細胞。在培養中,飼養層作為其他細胞的基底層并供給細胞因子,而它們自身不再生長或分裂(lim,j.w.andbodnar,a.,2002)。飼養層細胞的實例一般有人二倍體肺細胞、小鼠胚胎成纖維細胞、瑞士小鼠胚胎成纖維細胞,也可以是任何能夠供給對實現干細胞最佳的生長、活力和擴增有利的細胞成分和因子的有絲分裂期后細胞。在許多情況下,飼養細胞層不必使es細胞保持在未分化的、增殖性狀態下,因為白血病抑制性因子(lif)具有抗分化性質。因此,補充lif可用于將某些物種的mapc保持在未分化狀態下。
細胞可在低血清或無血清的培養基中培養。美國專利7,015,037記載了用于培養mapc的無血清培養基。已在低血清或無血清的培養基中培養了許多細胞。在這種情況下,在所述培養基中添加一種或多種生長因子。常用的生長因子包括但不限于成骨蛋白、堿性成纖維細胞生長因子、血小板衍生生長因子和表皮生長因子。參見例如美國專利no.7,169,610;7,109,032;7,037,721;6,617,161;6,617,159;6,372,210;6,224,860;6,037,174;5,908,782;5,766,951;5,397,706和4,657,866;所述全部專利都以引用的方式納入本文中以教導在無血清培養基中培養細胞。
培養中的細胞可維持在懸浮液中或貼壁于固體支持物,例如胞外基質成分。干細胞經常需要促進其附著于固體支持物的另外的因子,例如i型和ii型膠原、硫酸軟骨素、纖接蛋白、“超纖連蛋白(superfibronectin)”和纖接蛋白樣聚合物、明膠、聚-d-和聚-l-賴氨酸、血小板反應蛋白和玻璃粘連蛋白。本發明的一個實施方案利用纖連蛋白。參見,例如ohashietal.,naturemedicine,13:880-885(2007);matsumotoetal.,jbioscienceandbioengineering,105:350-354(2008);kirouacetal.,cellstemcell,3:369-381(2008);chuaetal.,biomaterials,26:2537-2547(2005);drobinskayaetal.,stemcells,26:2245-2256(2008);dvir-ginzbergetal.,fasebj,22:1440-1449(2008);turneretal.,jbiomedmaterrespartb:applbiomater,82b:156-168(2007);和miyazawaetal.,journalofgastroenterologyandhepatology,22:1959-1964(2007)。
細胞也可在"3d"(聚集的)培養中培養。一個實例為2008年1月18日提交的美國臨時專利申請no.61/022,121。
一旦在培養中形成,細胞可新鮮使用或冷凍使用,并使用例如含有40%fcs和10%dmso的dmem作為冷凍原種保存。本領域技術人員也可使用制備培養細胞的冷凍原種的其他方法。
藥物制劑
在某些實施方案中,純化的細胞群體存在于適應和適于送遞即生理上相容的組合物中。因此,所述干細胞群體的組合物通常還包括一種或多種緩沖劑(例如中性緩沖鹽水或磷酸緩沖鹽水)、碳水化合物(例如葡萄糖、甘露糖、蔗糖或葡聚糖)、甘露醇、蛋白、多肽或氨基酸例如甘氨酸、抗氧化劑、抑菌劑、螯合劑例如edta或谷胱甘肽、佐劑(例如氫氧化鋁)、使所述制劑與受者的血液相比等滲、低滲或稍微高滲的溶質、助懸劑、增稠劑和/或防腐劑。
在其他實施方案中,純化的細胞群體存在于適應或適于冷凍或保存的組合物中。
在許多實施方案中,用于給予受試者的細胞(或條件培養基)的純度為約100%。在其他實施方案中,其為95-100%。在某些實施方案中,其為85-95%。特別地,對于與其他細胞的混合物,所述百分比可為約10%-15%、15%-20%、20%-25%、25%-30%、30%-35%、35%-40%、40%-45%、45%-50%、60%-70%、70%-80%、80%-90%或90%-95%。或者,分離/純度可以細胞倍增的形式表示,其中細胞已發生例如10-20、20-30、30-40、40-50或更多次的細胞倍增。
給定體積的細胞數量可通過廣為人知的和常規的方法和儀器測定。給定體積的細胞混合物中的細胞的百分數可通過幾乎一樣的方法測定。可手動或通過使用自動細胞計數器容易地對細胞進行計數。給定體積的特定細胞可使用特異性染色和目測以及通過使用特異性結合試劑(一般為抗體)、熒光標記和熒光激活細胞分選器的自動化方法而測定。
倚賴多種因素選擇用于為給定用途給予所述細胞的制劑。在這些因素中,重要的是受試者的物種,待治療的病癥、功能障礙或疾病的性質及其在受試者中的狀態和分布,其他待給予的療法和試劑的性質,給藥的最佳途徑,經過該途徑的殘存性,給藥方案以及其他本領域技術人員明了的因素。具體地,例如,合適載體和其他添加劑的選擇會倚賴給藥的確切途徑和特定劑型的性質。
例如,細胞存活可為基于細胞的療法的效力的一個重要決定因素。這對于主要治療和輔助治療都是成立的。目標位點不適于細胞接種和細胞生長時會產生另一個問題。這可能會阻止治療性細胞接近該位點和/或移植到那里。本發明的多個實施方案包括增加細胞存活和/或克服因接種和/或生長屏障而引起的問題的措施。
細胞/培養基的水性懸液的最終配制一般包括將所述懸液的離子強度調節到等滲(即約0.1-0.2)以及將ph調節到生理ph(即約ph6.8-7.5)。所述最終制劑通常還含有液體潤滑劑例如麥芽糖,其必須為身體所耐受。示例性潤滑劑組分包括甘油、糖原、麥芽糖等。基于有機聚合物的材料,例如聚乙二醇和透明質酸以及非纖維狀膠原(優選琥珀酸化的膠原),也可用作潤滑劑。這些潤滑劑通常用于在注射位置上改善所注射生物材料的注射能力、潤透性和分散性,以及通過改變所述組合物的粘性來降低堵塞(spiking)的量。最后的制劑必然是可藥用載體中的細胞。
隨后將所述細胞置于注射器或其他注射裝置中以精確注射到組織缺損的位點上。術語“可注射”指所述制劑可在正常壓力和正常條件下以帶有低至25號針頭的注射器中給予,而沒有顯著堵塞。堵塞可導致組合物從注射器溢出而非注射到組織中。對于這一精確布置,需要細至27號(200μi.d.)、甚或30號(150μi.d.)的針頭。可通過這些針頭擠出的最大顆粒大小將因至少以下參數復雜變化:顆粒最大尺寸、顆粒長寬比(長:寬)、顆粒剛性、顆粒表面粗糙度和影響顆粒之間粘性的有關因素、懸浮液的粘彈性,以及通過針頭的流速。懸浮于牛頓流體中的剛性圓珠代表最簡單的情況,而在粘彈性流體中的纖維性或有分支的顆粒可能更復雜。
本發明的所述組合物的理想等滲性可使用氯化鈉或其他可藥用劑例如右旋糖、硼酸、酒石酸鈉、丙二醇、或其他無機或有機溶質實現。氯化鈉特別優選用于含有鈉離子的緩沖液。
如果需要,可使用可藥用的增稠劑將所述組合物的粘性保持在選定水平上。優選甲基纖維素,因為它可方便和經濟地獲得并且易于使用。其他合適的增稠劑包括例如黃原膠、羧甲基纖維素、羥丙基纖維素,卡波姆等。所述增稠劑的優選濃度取決于所選擇的試劑。重要的是使用實現選定粘度的量。粘性組合物通常是通過在溶液中加入所述增稠劑而制備的。
可藥用的防腐劑或穩定劑可用于增加細胞/培養基組合物的壽命。如果加入所述防腐劑,那么選擇不影響所述細胞活力或效力的組合物完全在本領域技術人員的能力范圍之內。
本領域技術人員會認識到所述組合物的組分在化學上應為惰性的。這在化學和藥學原理方面不對本領域技術人員構成問題。通過參考標準教科書,或者通過使用本公開內容、本文引用文獻和本領域通常可獲得的信息的簡單實驗(不涉及過多試驗)可以容易地避免問題。
無菌可注射溶液可通過以下方式制備:將用于實施本發明的細胞/培養基與不同量的其他所需成分摻入所需量的合適溶劑中。
在某些實施方案中,細胞/培養基被制成單位劑量的可注射形式,例如溶液、懸液或乳液。適于細胞/培養基注射的藥物制劑通常為無菌水性溶液和分散系。用于可注射制劑的載體可為含有如下物質的溶劑或分散介質:例如,水、鹽水、磷酸緩沖鹽水、多元醇(例如甘油、丙二醇、液體聚乙二醇等)以及它們的合適混合物。
本領域技術人員可容易地確定將在本發明方法中給予的組合物中的細胞和可選添加劑、媒介(vehicle)和/或載體的量。通常,任何添加劑(除所述細胞以外)都以0.001-50重量%的量存在于溶液例如磷酸緩沖鹽水中。所述活性組分以微克至毫克量級存在,例如約0.0001到約5重量%,優選約0.0001到約1重量%,最優選約0.0001到約0.05重量%,或者約0.001到約20重量%,優選約0.01到約10重量%,最優選約0.05到約5重量%。
在某些實施方案中,將細胞封裝(膠囊化)給藥,特別是當膠囊化可提高治療效力時或者提供處理和/或貯藏壽命上的優點時。在某些實施方案中,當膠囊化增加細胞介導的免疫抑制的效力時,它也因此降低對免疫抑制藥物治療的需要。
此外,在某些實施方案中,膠囊化提供可進一步降低受試者對所述細胞(通常在同種異體移植中具有免疫原性或者只有微弱免疫原性)的免疫反應的針對受試者免疫系統的屏障,從而減輕由于給予所述細胞而可能發生的任何移植排斥或炎癥。
細胞可在植入前通過膜以及膠囊進行封裝。應考慮到,可使用可用于細胞膠囊化的多種方法中的任意一種。在某些實施方案中,細胞被單獨封裝。在某些實施方案中,許多細胞被封裝在同一膜內。在其中所述細胞在植入后被取出的實施方案中,將許多細胞封裝在例如單一膜內的較大尺寸的結構可提供方便的取出方法。
在微囊化細胞的多個實施方案中可使用多種材料。這些材料包括例如聚合物膠囊,藻酸鹽-聚-l-賴氨酸-藻酸鹽微膠囊、聚-l-賴氨酸藻酸鋇膠囊、藻酸鋇膠囊、聚丙烯腈/聚氯乙烯(pan/pvc)中空纖維以及聚醚砜(pes)中空纖維。
可用于細胞給藥的細胞微囊化技術為本領域技術人員已知,在例如chang,p.,etal.,1999;matthew,h.w.,etal.,1991;yanagi,k.,etal.,1989;caiz.h.,etal.,1988;chang,t.m.,1992以及美國專利no.5,639,275(其描述了例如用于長期維持穩定地表達生物活性分子的細胞的生物相容膠囊)中描述了這些技術。膠囊化的其他方法見歐洲專利公開no.301,777和美國專利no.4,353,888;4,744,933;4,749,620;4,814,274;5,084,350;5,089,272;5,578,442;5,639,275和5,676,943。所有上述文獻與細胞膠囊化有關內容都以引用的方式納入本文。
某些實施方案將細胞加入聚合物中,例如生物聚合物或合成聚合物。生物聚合物的實例包括但不限于纖連蛋白、纖維蛋白(fibin)、纖維蛋白原(fibrinogen)、凝血酶、膠原和蛋白聚糖。其他因子例如上面討論的細胞因子也可被加入所述聚合物中。在本發明的其他實施方案中,細胞可被加入三維凝膠的間隙中。大的聚合物或凝膠通常通過手術植入。可配制成足夠小的顆粒或纖維的聚合物或凝膠可通過其他常規的、更方便的、非手術的途徑給予。
給藥
組合物可根據諸如具體患者的年齡、性別、體重和病癥,以及將給藥的制劑(例如固體對液體)等因素,按劑量并通過醫藥和獸醫領域技術人員公知的技術給藥。人類或其他哺乳動物的劑量可由技術人員根據本公開內容、本文引用的文獻以及本領域的知識,不需過度實驗而確定。
適用于本發明多個實施方案的細胞/培養基的劑量取決于多種因素。其對于不同的情況可發生相當大的變動。確定主要和輔助治療的最佳給藥劑量的參數一般包括下面的一些或全部:要治療的疾病及其階段;受試者的物種、它們的健康、性別、年齡、體重和代謝速率;受試者的免疫活性;其他正在給予的療法;以及根據受試者的歷史或基因型預測的可能并發癥。所述參數還可包括:所述細胞是同基因的、自體的、同種異體的還是異種的;它們的效能(具體活性);使所述細胞/培養基有效而必須靶向的位點和/或分布;以及所述位點的這些特性例如細胞/培養基的可達性和/或細胞的植入性。其他參數包括與其他因子(例如生長因子和細胞因子)的共給藥。給定情況下的最佳劑量還要考慮將細胞/培養基配制的方式、給予它們的方式、給藥后細胞/培養基定位到靶標位點的程度。最后,最佳劑量的確定必須提供這樣的有效劑量,即所述劑量既不低于最大有益效果的閾值也不高于劑量相關的有害作用超過增加劑量的益處的閾值。
對于某些實施方案,細胞的最佳劑量處在用于自體、單核骨髓移植的劑量范圍內。對于非常純的細胞制劑,在不同實施方案中,每次給藥的最佳劑量可為104-108細胞/kg接受者質量。在某些實施方案中,每次給藥的最佳劑量為105-107細胞/kg。在許多實施方案中,每次給藥的最佳劑量為5×105-5×106細胞/kg。用于參考,前述較高的劑量與用于自體單核骨髓移植的有核細胞的劑量相似。某些較低的劑量與用于自體單核骨髓移植的cd34+細胞/kg的數量相似。
應理解,單份劑量可一次全部、分份或在一段時間內連續遞送。也可將整份劑量遞送到單個位置或分份分散到若干個位置。
在各個實施方案中,細胞/培養基可以以初始劑量給藥,然后通過后續給藥保持。細胞/培養基可在開始通過一種方法給藥,然后通過相同方法或者一種或多種不同方法給藥。水平可通過繼續給予所述細胞/培養基而保持。各個實施方案通過靜脈內注射在開始時給予所述細胞/培養基和/或保持它們在受試者中的水平。在多個實施方案中,可根據患者的病情和本文其他地方討論的其他因素使用其他給藥形式。
應注意,人類受試者接受治療的時間通常長于實驗動物,然而,治療的時間通常與疾病進展的時間和治療效果成比例。本領域技術人員在使用在人和/或動物(諸如大鼠、小鼠、非人靈長類等)中實行的其他方法的結果時會考慮這一因素,以確定用于人的合適劑量。基于這些考慮并根據本公開內容和現有技術所提供的指導,這種確定可使技術人員不需過度實驗而確定劑量。
初始給藥和后續給藥或連續給藥的合適方案可以都相同或者可以是變化的。合適方案可由技術人員根據本公開內容、本文引用的文獻以及本領域的知識確定。
治療的劑量、頻率和持續時間取決于許多因素,包括疾病的性質、受試者以及可能給予的其他療法。因此,多種方案可用于給予所述細胞/培養基。
在某些實施方案中,將細胞/培養基以一個劑量給予受試者。在其他一些實施方案中,將細胞/培養基以一系列的兩個劑量或多個劑量連續給予受試者。在其中以單個劑量、兩個劑量和/或多于兩個的劑量給予細胞/培養基的其他一些實施方案中,劑量可相同或不同,并且給藥之間的間隔可相同或不同。
細胞/培養基可在各種不同時間內以多個頻率給藥。在某些實施方案中,細胞在小于1天的時間內給藥。在其他實施方案中,它們在2、3、4、5、或6天的時間內給藥。在某些實施方案中,細胞在數周的時間內以每周一次或多次給藥。在其他實施方案中,細胞在數周的時間內給藥持續一到數月。在多個實施方案中,細胞可在數月的時間內給藥。在其他實施方案中,細胞可在一或數年的時間內給藥。通常治療時間是與疾病進展的時間、所用療法的效果以及要治療的受試者的情況和反應成比例的。
例如,本文所解釋的脊髓損傷可分兩個階段。在動物模型中,在第一階段,巨噬細胞不滲入所述損傷,此階段持續約24小時。然而,在第二階段,巨噬細胞滲入所述損傷,當評定治療方案時可考慮此事件后果。在一個實施方案中,如果預期會與營養不良的軸突相互作用的巨噬細胞或其他細胞會滲入,則甚至在第一階段過程中即給予細胞,例如在剛損傷后或與損傷盡可能接近。然后,可繼續治療以與巨噬細胞的初始和后續滲入同步,并且可預防性地繼續治療,或者在確定巨噬細胞或其他相關細胞不再滲入所述損傷時可能中斷治療。
實施例
實施例i
"glialscarmodel"aggrecan-lamininopposingspotgradients(tometal..2004;steinmetzetal..2005)。這些參考文獻以引用的方式納入以教導神經膠質疤痕模型。此模型提供了細胞、蛋白、培養基等對在體外降低粘附/回縮的有效性的測定法。
如果抑制性基質存在于比體內損傷后形成的空間組織更相似的空間組織中,那么pg可誘導軸突中所謂的營養不良狀態。為此,pg聚集蛋白聚糖和生長促進分子層粘連蛋白的溶液的斑點被置于硝酸纖維素蓋玻片上并空氣干燥。
干燥的連續人工產物形成了粗略的梯度,其中所述斑點的邊緣與中心相比含有越來越高的聚集蛋白聚糖濃度。所述邊緣的最外部與任何更接近中心的區域相比含有更低的層粘連蛋白的濃度。最佳ecm濃度(0.7mg/ml聚集蛋白聚糖和5μg/ml層粘連蛋白)導致了良好的細胞貼附。因此,高聚集蛋白聚糖-低層粘連蛋白的外邊緣似乎是對再生神經突特別不利的區域。沒有神經突通過穿過斑點清晰的外邊界從環繞的層粘連蛋白進入所述斑點。從所述斑點的中心內向心生長的纖維能夠進入所述邊緣的內部,但不能再向前生長。一旦進入所述梯度,軸突就顯得被困住。在所述梯度內,在神經突末端處形成了球棒狀的、營養不良的端球。為了觀察所述“營養不良的末端”的表現,使用了縮時顯微術(time-lapsemicroscopy)。營養不良的生長錐經常努力前進較短的距離,但不可避免的,所述艱難行進的生長錐會聚攏成更致密的球并回縮,只是為了再次開始移動。
實施例ii
軸突回縮和巨噬細胞
總述
在體內,在背柱破碎脊髓損傷后,在位于切斷的軸突末端的營養不良的回縮球棒狀物與ed-1+細胞之間發現了緊密關聯(圖3)。使用了神經膠質疤痕的體外模型(tometal.,2004;steinmetzetal.,2005)來實時檢查軸突與ed-1+細胞之間的相互作用。營養不良的生長錐與ed-1+巨噬細胞之間直接的細胞-細胞接觸誘導了長距離軸突回縮(圖4、5)。使用氯膦酸鹽脂質體(popovichetal.,1999)在體內清除巨噬細胞的結果是經氯膦酸鹽處理的動物與對照相比軸突回縮的明顯減少。這些數據表明ed-1+細胞通過物理的細胞-細胞相互作用直接引起受損傷脊髓軸突的回縮。
結果
1.脊髓損傷后上行脊柱感覺軸突大幅回縮
發明人認為活化的巨噬細胞的滲入能夠在軸突回縮中起直接作用。在亞急性和慢性脊髓損傷中,在活化的巨噬細胞和再生軸突的末端之間發現了緊密關聯,使得這兩種細胞類型之間可發生直接物理相互作用。然后表征背柱破碎損傷后感覺軸突回縮的程度并且將其與巨噬細胞滲入到損傷中相關。將成體雌性sprague-dawley大鼠的脊柱在節段c8打碎,并通過dextran-texasred標記坐骨神經追蹤受損傷神經元子群。在損傷后2、4、7、14和28天收集脊髓組織并測量標記纖維末端與損傷中心之間的距離。到損傷后2天,軸突已經回縮343±46.92um的平均距離(平均值±標準偏差),然而,此早期回縮最可能是因為神經元自身內的固有機制(kerschensteineretal.,2005)。需要強調的是,在損傷后2天,損傷主要由有活性的星形細胞(gfap+)和少數ed-1+細胞構成,所述ed-1+細胞最可能是原有的小神經膠質。在損傷后2天和7天之間損傷內的ed-1+細胞的數量急劇增加,其大部分是最可能滲入的巨噬細胞(popovichetal.,1997;donnellyandpopovich,2007)。脊柱內上行感覺纖維的第二階段回縮在第一周最快地出現,然后在隨后的幾周逐步地出現。到損傷后28天,軸突已從損傷中心回縮774±70.26um的平均距離。這些數據表明上行感覺軸突回縮的時間與損傷內ed-1+細胞的滲入和集聚在空間和時間上一致。
2.活化的巨噬細胞的清除減少體內軸突回縮
體內軸突回縮的第二階段主要在損傷后2天和7天之間——與巨噬細胞滲入的時間一致——發生在上行脊柱感覺軸突中。為進一步弄清巨噬細胞在體內軸突回縮中的作用,用氯膦酸鹽脂質體處理動物以清除循環的單核細胞/巨噬細胞(vanrooijenetal.,1997;popovichetal.,1999)。動物每隔一天接受氯膦酸鹽脂質體注射,在受傷前一天開始處理,以清除循環的單核細胞/巨噬細胞。然后在損傷后2天、4天和7天對動物評定軸突回縮(圖3)。與接受對照脂質體的動物在損傷后4天和7天的回縮(分別為586±42.89um和806±62.71um)比較,接受氯膦酸鹽脂質體注射的動物在損傷后4天和7天表現出回縮的明顯減少(分別為402±81.85um和439±46.33um)。回縮減少與經氯膦酸鹽處理的動物與接受空脂質體對照的動物相比損傷內ed-1+細胞數量的顯著減少相關。氯膦酸鹽脂質體處理還導致損傷中心的gfap+星形細胞突起(process)的增加,與之前觀察到的巨噬細胞清除導致空穴形成減少的現象一致(popovichetal.,1999)。重要的是,在損傷后2天,經氯膦酸鹽處理和經對照脂質體處理的動物中顯現的回縮量沒有差別。此時尚未出現巨噬細胞滲入,表明軸突回縮的第一階段是不巨噬細胞依賴的,可能是由于內生神經元機制或者可能是由于與活化的原有小神經膠質的相互作用。氯膦酸鹽介導的循環巨噬細胞/單核細胞的清除阻止了通常在損傷后4天和7天出現的軸突回縮,表明回縮的第二階段是由滲入巨噬細胞的作用導致的。在經氯膦酸鹽處理的動物中未出現明顯的再生(即軸突延長至損傷中心之外)。
3.在神經膠質疤痕的體外模型中營養不良的生長錐在與巨噬細胞接觸后大幅回縮
在體內觀察到ed-1+細胞與受損傷的軸突的緊密關聯表明這些細胞之間的相互作用可能在軸突回縮中起作用。研究了神經膠質疤痕的體外模型中成體感覺神經元軸突與巨噬細胞之間的相互作用。在30分鐘的基線觀察期后,將nr8383巨噬細胞加入培養物中并監測它們與營養不良的軸突的相互作用。在營養不良的軸突與巨噬細胞之間經常觀察到直接的細胞-細胞接觸。這些接觸持續較長一段時間,并且當與巨噬細胞的遷移結合時,引起對軸突的強烈操控,導致軸突有力地彎曲并從基底抬起。很明顯,強的、長時間的附著可在所述兩種細胞類型之間發生,因為在巨噬細胞移開軸突之后經常仍然存在連接所述兩種細胞的長拖尾突起。然而,巨噬細胞誘導的回縮不永久阻止軸突的再生長,因為某些在回縮后不再與巨噬細胞接觸的軸突能夠延長直至這些軸突再度營養不良。這兩種細胞類型之間直接的細胞-細胞接觸最終導致了軸突在全部接觸時間的大幅回縮。因此,在神經膠質疤痕的體外模型中巨噬細胞接觸誘導了營養不良的軸突的回縮。
4.直接的物理的細胞-細胞相互作用對于巨噬細胞誘導的回縮是必需的
有多次觀察到巨噬細胞移動至非常接近但未接觸營養不良的軸突處的情況,在這些情況下未觀察到軸突回縮。為了確定軸突與巨噬細胞之間的物理相互作用是否是誘導軸突回縮所必需的或者巨噬細胞產生的因子是否足夠,用胰蛋白酶處理巨噬細胞以除去胞外蛋白,然后將其加至drg培養物。用胰蛋白酶對巨噬細胞的預處理不妨礙巨噬細胞的大幅遷移和與軸突的多次碰撞。然而,所述處理完全阻止了巨噬細胞物理附著于軸突,因此在不存在長時間的直接細胞-細胞接觸的情況下未觀察到回縮。然而,巨噬細胞分泌誘導回縮的因子是可能的。為檢測這一假設,在體外將巨噬細胞條件培養基加至營養不良的軸突。巨噬細胞條件培養基不誘導回縮。因此,在不存在與營養不良的軸突的物理相互作用的情況下,在軸突附近僅存在巨噬細胞或其分泌的因子均不能誘導軸突回縮。
5.巨噬細胞誘導的回縮中基底的作用
為測定基底在軸突回縮中是否起作用,在均勻生長促進層粘連蛋白基底上培養成體感覺神經元,所述基底不產生營養不良的生長錐。借助多個絲狀偽足(filopodia)和板層形偽足(lamellapodia)使層粘連蛋白上的生長錐變平,并且全部軸突延伸均以恒定速度發生。當將巨噬細胞加至這些培養物時,觀察到了與軸突的直接細胞-細胞接觸。然而,這些接觸是短暫的并很快被破壞,幅度不那么大,并且未導致軸突回縮。膜接觸點的殘留部分被快速重吸收回神經元中,并且生長錐繼續延伸穿過無阻礙的基底。因此,巨噬細胞誘導的軸突回縮是基底依賴的;在允許的基底層粘連蛋白上活動生長狀態的神經元對巨噬細胞接觸不敏感,與cspg梯度誘導的營養不良狀態下的神經元不同。
6.活化的原代巨噬細胞也誘導軸突回縮
另一問題是,原代巨噬細胞在體外是否以與nr8383巨噬細胞系同樣的方式與營養不良的軸突相互作用。收集成年sprague-dawley大鼠骨髓中的祖細胞并使其在體外分化成巨噬細胞,得到多于80%的ed-1+細胞的培養物。此具體巨噬細胞群體已顯示出保留有在脊髓損傷中出現的巨噬細胞的表型、形態和功能特征,而不同于從其他身體來源收集的群體(longbrakeetal.,2007)。然后測定了原代巨噬細胞誘導軸突回縮的能力。未刺激的原代巨噬細胞不能誘導回縮。當加至斑點梯度神經元培養物時,這些巨噬細胞附著于基底但不能動,呈現休眠狀態的巨噬細胞特征。僅當將巨噬細胞直接置于營養不良的軸突上時才出現與軸突的接觸。這些巨噬細胞及其細胞-細胞相互作用均未表現出之前用細胞系巨噬細胞觀察到的任何物理特征,即無牽拉、無經細胞突出的物理附著的跡象等。
巨噬細胞可能必須處于活動狀態以與營養不良的軸突相互作用。在培養中用活化細胞因子干擾素γ刺激原代巨噬細胞,然而加至縮時培養皿中。盡管這些巨噬細胞呈現中度活化狀態和偏圓的形態,然而它們仍然不能動并且不與營養不良的軸突形成強附著,因此不導致軸突回縮。再用干擾素γ和脂多糖(lps)的結合物刺激原代巨噬細胞,然后加至drg培養物中。這些巨噬細胞表現出活化的巨噬細胞的形態和表現:圓的、噬菌細胞的形狀和高能動性。這些活化的巨噬細胞以與細胞系巨噬細胞同樣的頻率誘導營養不良的軸突回縮。它們顯示出與營養不良的軸突的有力物理相互作用,導致細胞之間的強附著以及軸突的物理抓附、牽拉和從基底的抬起。當處于活化狀態時,原代巨噬細胞在體外誘導了營養不良的軸突的回縮,認可了在此體外軸突回縮的研究中使用細胞系巨噬細胞。因此,實驗主要用nr8383巨噬細胞系進行,因為該細胞系形成了處于恒定活化狀態的純細胞群體,其類似于在脊髓損傷內發現的沒有額外刺激的巨噬細胞。
7.活化的小神經膠質在體外能夠中度誘導軸突回縮
盡管巨噬細胞通常在損傷后3天之前不會侵入受損傷的脊髓,然而cns內原有的小神經膠質會立即對損傷作出反應(watanabeetal.,1999)。損傷內的小神經膠呈現活化和噬菌細胞的狀態,很像活化的巨噬細胞。在損傷后2天,在典型的巨噬細胞滲入之前,在損傷內發現有限數量的ed-1+細胞,這些細胞最可能是原有的小神經膠質。使用體外模型評定了小神經膠質對軸突回縮的可能作用。從p1sprague-dawley大鼠收集了腦皮質小神經膠質并使其在體外成熟,然后將其加入縮時培養物中。類似于原代巨噬細胞,在培養中必須用干擾素γ和lpc刺激原代小神經膠質以使其被活化。未被刺激的小神經膠質不能附著于層粘連蛋白/集聚蛋白聚糖斑點梯度基底,這阻止所述細胞在本發明人的模型中與營養不良的軸突相互作用。然而,被刺激的小神經膠質附著并與軸突物理相互作用,在50%的接觸時間內誘導回縮,然而,活化的小神經膠質與營養不良的軸突之間的接觸不如巨噬細胞的強。因此,實驗活化的小神經膠質也可在軸突回縮的誘導中起作用。
8.另一種cns細胞類型——星形細胞——不能在體外誘導軸突回縮
另一個問題是,在體外誘導營養不良的軸突的回縮是否是對在損傷的脊髓內常見的噬菌細胞類型特異的,而不僅是營養不良的軸突與任何其他細胞類型的相互作用的結果。星形細胞是cns的損傷后的神經膠質疤痕的一個不可或缺的組分。它們大量存在并廣泛接觸再生的軸突。使腦皮質星形細胞在體外成熟,然后加至drg培養物中。星形細胞附著于基底并廣泛接觸營養不良的軸突。一旦與基底結合,星形細胞就沿集聚蛋白聚糖梯度迅速遷移,遠離邊緣。星形細胞突起分布在軸突上,有時導致軸突的側向位移。然而,這些接觸不導致被接觸的軸突回縮。因此,回縮的誘導對與ed-1+噬菌細胞的相互作用是特異的,而不僅是與任何其他細胞類型的物理相互作用。
材料和方法
1.背柱破碎損傷模型
使用33只成年雌性sprague-dawley大鼠(250-300g)進行體內研究。對于所有手術步驟,均用吸入的異氟醚氣(2%)麻醉大鼠。進行t1椎板切除術來暴露c8脊髓節的背面。用30號針頭在距離中線0.75mm的兩側進行硬腦膜切開術。然后通過將dumont#3jeweler鑷子在c8處插入背脊至1.0mm的深度并壓住鑷子,保持壓力10秒鐘并再重復2次來制造背柱破碎損傷。通過觀察到白色物質的清除來確認損傷的完成。然后用凝膠膜覆蓋硬腦膜上的洞。用4-0尼龍線縫合肌肉層,然后用外科封縫釘閉合皮膚。在閉合切口后,動物沿切口皮下接受麻卡因(marcaine,1.0mg/kg)并肌內注射丁丙諾啡(0.1mg/kg)。手術后,在麻醉恢復期間用加熱燈使動物保暖,并使其自由取食和飲水。將動物在損傷后2、4、7、14或28天處死(每組n=3)。所有動物操作均依照casewesternreserveuniversity的animalresourcecenter的指南和方案進行。
2.巨噬細胞清除
動物在背柱破碎損傷前的一天接受脂質體封裝的氯膦酸鹽或空脂質體對照的腹膜內注射,并且還在損傷后一天(對于2d時間點)、損傷后1天和3天(對于4d時間點)和損傷后1、3和5天(對于7d時間點)接受上述腹膜內注射(每組n=3)。氯膦酸鹽由rochediagnosticsgmbh,mannheim,germany贈予。氯膦酸鹽如現有技術所述封裝在脂質體中(vanrooijenandsanders,1994)。
3.軸突標記
處死前2天,用texas-red綴合的3000mw葡聚糖單側標記脊柱。簡言之,暴露右后肢的坐骨神經,用dumont#3鑷子擠壓10秒鐘并再重復兩次。在擠壓位點處用hamilton注射器將1.0ul3000mw葡聚糖-texas-red的10%無菌水溶液注射到坐骨神經中。用4-0尼龍線縫合肌肉層,然后用外科封縫釘閉合皮膚。在閉合切口后,動物沿切口皮下接受麻卡因(1.0mg/kg)并肌內注射丁丙諾啡(0.1mg/kg)。手術后,在麻醉恢復期間用加熱燈使動物保暖,并使其自由取食和飲水。將動物在標記后2天用過量異氟醚處死并用pbs繼而用4%pfa灌注。收集組織并在4%pfa中后固定并進行免疫組織化學測定。
4.免疫組織化學測定
將組織在4%pfa后固定過夜,然后浸沒在30%蔗糖中過夜,在oct封裝介質中冷凍,然后在恒冷切片機上切成20um的縱向切片。然后將組織分別用抗gfap(accuratechemicalandscientificcorporation,westbury,ny)、抗ed-1(millipore,billerica,ma)染色,并分別用alexafluor-405或alexafluor-488(invitrogen,carlsbad,ca)孵育,然后在zeissaxiovert510激光掃描共焦顯微鏡上成像。
5.體內軸突回縮定量
對每只動物分析了始于每只動物的脊髓背面下200um深處的三個連續切片以定量軸突回縮。通過特征性gfap和ed-1染色模式和標記的軸突末端之間的距離確定損傷中心,并使用zeisslsm5imagebrowser軟件確定中心位置。將一組中所有動物的所有切片的測量值取平均以得出每個時間點的回縮平均距離。
用于追蹤受損傷纖維的技術標記位于脊柱內非常淺處的軸突。同時,標記的纖維的數量可由于注射示蹤劑的節段的坐骨神經的肌纖維自發性收縮的程度而變化。出于多種原因,僅定量在該深度的標記的軸突。此深度在所有動物中均含有標記的纖維,而某些動物在更深的深度不具有標記的纖維。損傷的線性程度在所述脊柱的更深節段增加。因此位于脊髓更深處的軸突受到了比在更淺節段的損傷大很多的損傷。回縮距離的定量必須發生損傷的類似位置以允許動物和組之間的準確比較。對標記軸突的整個群體的定量可因標記程度的不同而導致結果的偏差。相反,對標記軸突的具體群體和位置進行定量,從而能夠在所有動物中對它們進行一致的檢查和準確的定量。
6.drg解離
如現有技術所述收集drg(tometal.,2004;daviesetal.,1999)。簡言之,從成體雌性sprague-dawley大鼠切下drg(zivicmiller,harlan)。除去中心根和外周根,并在膠原酶ii(200u/ml,worthington)和dispaseii(2.5u/ml,roche)的hbss溶液中孵育神經節。將消化的drg沖洗并在新鮮hbss-cmf中輕輕研磨三次,然后低速離心。然后將解離的drg重懸浮在補充有b-27、glutamax、青霉素/鏈霉素(均來自invitrogen)的neurobasal-a培養基中并計數。將drg以3000個細胞/ml的密度置于delta-t培養皿上(fisher,pittsburgh,pa),共6000個細胞/培養皿。
7.縮時培養皿制備
與tometal.,2004所述類似地制備delta-t細胞培養皿(fisher,pittsburgh,pa)。簡言之,用2號鉆頭通過每個培養皿的上半部鉆一個洞,以形成在縮時顯微術過程中向培養物中加細胞、酶、抑制劑等的進口。然后,將培養皿用無菌水沖洗并在室溫下以聚-l-賴氨酸(0.1mg/ml,sigma)包被過夜,然后用無菌水沖洗并使其干燥。通過吸取2.0ul的聚集蛋白聚糖溶液(2.0mg/ml,sigma,溶于hbss-cmf,invitrogen)于培養物表面上并使其干燥,形成聚集蛋白聚糖梯度斑點。每個培養皿中放置6個斑點。在聚集蛋白聚糖斑點完全干燥后,將培養皿的整個表面在層粘連蛋白的hbss-cmf溶液(10ug/ml,bti,stoughton,ma)中于37℃溫浴3小時。然后撤除層粘連蛋白浴,立即將細胞鋪板。只用層粘連蛋白浴不用聚集蛋白聚糖如上所述制備了只含有層粘連蛋白基底的培養皿。此處所用的基底的濃度不同于tometal.,2004中所用的濃度。顯微術的清晰度可通過去除硝酸纖維素而從培養皿制備方案改善。然而,為補償結合于培養皿表面的基底的不同,將所用的基底濃度重新校準為上面所列的濃度。
縮時成像后,將drg在4%pfa中固定并用抗iii型b-微管蛋白(1:500;sigma,st.louis,mo)和抗硫酸軟骨素(cs-56,1:500,sigma)免疫染色。
8.細胞系巨噬細胞培養物
如yinetal.(2003)所述培養成體sprague-dawley肺泡巨噬細胞系nr8383細胞(atcc#crl-2192)。簡言之,在未包被的組織培養燒瓶(corning)中,將細胞在補充有15%fbs、glutamax、penn/strep(invitrogen)和碳酸氫鈉(sigma)的f-12k培養基(invitrogen)中培養,并每周加料2-3次。此細胞系形成貼壁和懸浮細胞的混合培養物,并在加料時通過收集和重鋪板懸浮細胞進行傳代。為制備用于縮時顯微術實驗的細胞系巨噬細胞,將細胞用0.5%胰蛋白酶/edta(sigma)收集并用無血清f-12k洗滌3次,然后以1.0×106/ml的密度鋪板于含無血清f-12k的未包被組織培養燒瓶中。在用于縮時實驗前的1天,用edta和細胞刮刀收集培養的細胞系巨噬細胞并以2.5×105/70ul的密度重懸浮于添加有hepes(50um,sigma)的如上述補充neurobasal-a中。
9.原代骨髓源巨噬細胞培養物
基于現有技術方案(tobianetal.,2004)收集骨髓祖細胞。簡言之,將股骨從成年雌性sprague-dawley大鼠(225-275g,harlan)取下。除去股骨末端,將含有補充有10%fbs、glutamax、penn/strep、β-巰基乙醇和hepes(invitrogen)(d10f)的冷dmem的注射器插入股骨,沖出骨髓并收集。然后使所得細胞混合物通過70微米的濾器并離心。然后除去上清液,將所得細胞片狀沉淀物重懸浮于akt裂解緩沖液(biowhitacre)中以裂解紅細胞,然后離心。除去上清液并將含有骨髓前體細胞的片狀沉淀物重懸浮并鋪板于另補充有20%ladmac細胞系條件培養基(cliffordharding贈予)的上述dmem中以誘導分為成巨噬細胞。在培養第5、7、9天對細胞加料,在第10天收集細胞用于實驗。縮時實驗前的1天,用胰蛋白酶/edta收集原代巨噬細胞,用d10f洗滌3次并以1.0×106/ml的密度鋪板于含d10f的未包被陪替氏培養皿(falcon)中。第2天,用edta和細胞刮刀收集原代巨噬細胞并以5.0×105/70ul的密度重懸浮于加hepes的neurobasal-a中,用于縮時顯微實驗。
10.腦皮質星形細胞制備
通過取出p0-p1大鼠的腦皮質,切碎并用0.5%胰蛋白酶的edta溶液處理來收集腦皮質星形細胞。將細胞接種于以多聚l-賴氨酸包被的t75燒瓶上的含有10%fbs(sigma)和2mmglutamax的dmem/f12(invitrogen)中,并搖動,4小時后除去未貼壁的細胞。星形細胞在培養中成熟至少28天。用edta和細胞刮刀收集星形細胞并以5.0×105/70ul的密度重懸浮于加hepes的neurobasal-a中,用于縮時顯微實驗。
11.皮質小神經膠質制備
通過取出p0-p1大鼠的腦皮質,切碎并用0.5%胰蛋白酶的edta溶液處理來收集腦皮質小神經膠質。將細胞鋪板于以多聚l-賴氨酸包被的t75燒瓶上的含有20%fbs(sigma)和2mmglutamax的dmem/f12(invitrogen)中,維持5-7天。縮時實驗前的1天,搖動燒瓶以除去貼壁不牢的細胞并將這些細胞以1.0×106/ml的密度鋪板于含d10f的未包被陪替氏培養皿(falcon)中。第2天,用edta和細胞刮刀收集原代小神經膠質并以5.0×105/70ul的密度重懸浮于加hepes的neurobasal-a中,用于縮時顯微實驗。
12.縮時顯微術研究
將drg神經元于37℃孵育48小時,然后進行縮時成像。將含有hepes(50um,sigma)的neurobasal-a培養基加至培養物中,然后轉至加熱的臺架儀器中。使用zeissaxiovert405m顯微鏡和100×油浸物鏡,每30秒取得縮時成像,持續3小時。選擇徑直延伸到斑點邊緣并具有特征性營養不良形態的生長錐。觀察神經元30分鐘以確定基線表現,然后加入其他細胞類型(原代巨噬細胞n=3,其他組n=6)。加入細胞后,觀察生長錐150分鐘。本發明人用metamorph軟件追蹤了延伸/回縮和生長速率。
13.統計學分析
用minitab15軟件通過單因素或雙因素anova或普通線性模型(如果適用)以及tukey事后檢驗分析數據。
討論
這些結果首次表明了巨噬細胞可通過直接的物理接觸誘導營養不良的成體軸突回縮的確鑿證據。回縮的誘導同時倚賴神經元的生長狀態和巨噬細胞的活化狀態。在體外,原代骨髓源巨噬細胞必須用干擾素γ和lps刺激以達到與細胞系巨噬細胞和脊髓損傷內的巨噬細胞誘導軸突回縮類似的活化狀態。這說明巨噬細胞的體內表現是狀態依賴的并與之前表明巨噬細胞滲入僅在髓磷脂變性情況下與軸突回縮相關的研究(mcphailetal.,2004)是一致的。本研究表明成體感覺神經元僅在處于由抑制性cspg梯度誘導的停止生長的營養不良狀態時才對巨噬細胞誘導的回縮敏感。在均勻層粘連蛋白基底上處于生長活化狀態的成體神經元迅速中斷與巨噬細胞的接觸并且不發生回縮。
回縮的誘導可能包括多種內部和外部機制。本研究表明如果巨噬細胞與營養不良的軸突之間無直接的細胞-細胞接觸則不發生回縮。僅加入經胰蛋白酶處理的巨噬細胞或巨噬細胞條件培養基不足以誘導回縮。巨噬細胞無需特別接觸在生長錐處的營養不良的軸突來誘導回縮。與活化的巨噬細胞接觸可能觸發軸突內遠離其營養不良末端的信號傳遞通路。
存在若干候選結合配偶體,通過所述配偶體巨噬細胞和神經元可物理地相互識別并相互作用。巨噬細胞可使用αv和β1整合素受體來識別和結合軸突玻璃粘連蛋白(sobeletal.,1995),并且巨噬細胞與變性周圍神經的附著通過阻斷β整合素被部分地減弱(brownetal.,1997)。視神經損傷后,軸突表達ephrinb3,其被巨噬細胞上的ephb3受體識別(liuetal.,2006)。唾液酸粘附素——一種巨噬細胞特異的唾液酸受體——存在于神經元細胞膜上(kelmetal.,1994;tangetal.,1997)。巨噬細胞還識別暴露在發生細胞凋亡的細胞外膜表面上的磷脂酰絲氨酸(deetal.,2002;desimoneetal.,2004),所述磷脂酰絲氨酸可標示受損傷的神經元以進行內吞作用。此外,fractalkine是主要在cns神經元的表面上表達的趨化因子,而其受體cx3cr1存在于巨噬細胞上(zujovicetal.,2000;umeharaetal.,2001)。必須進行進一步的研究來確定,這些分子中的哪些(如果有的話)在營養不良的成體神經元表面上表達或上調,確定它們為巨噬細胞識別的靶標。
本研究證實了上行脊柱通路發生回縮(borgensetal.,1986;mcphailetal.,2004;stirlingetal.,2004;bakerandhagg,2005;bakeretal.,2007)并且時間進程與巨噬細胞的滲入一致。軸突回縮已在脊髓內的其他通路包括下行皮質脊髓通路(fishmanandkelley,1984;iizukaetal.,1987;hilletal.,2001;seifetal.,2007)、延髓通路(houleandjin,2001)和紅核脊髓通路(schwartzetal.,2005;caoetal.,2007)中被檢測。重要的是認為軸突回縮分為兩個不同的階段。對成年小鼠神經軸索斷裂的上行感覺軸突進行體內成像的最近研究表明,損傷前幾個小時內軸突回縮約300um,然后軸突在損傷后前3天中穩定(kerschensteineretal.,2005)。因此,本研究的焦點是軸突回縮的第二、后一階段,這是因為活化的巨噬細胞而不是因為所述神經元的固有性質。
本研究表明,在損傷之前以及整個正常回縮期間用氯膦酸鹽脂質體處理阻止了損傷后2天至7天之間的軸突回縮的第二階段。以前的工作已經表明,氯膦酸鹽脂質體介導的巨噬細胞清除導致損傷體積減小和神經元存活增加(popovichetal.,1999;vanrooijenandvankesteren-hendrikx,2002)。盡管這些作者強調巨噬細胞清除對軸突再生的作用,然而本研究的數據表明經氯膦酸鹽脂質體處理的動物的損傷的軸突含量增加可能(至少部分)是由于軸突回縮變弱所致。
實施例iii
干細胞可阻止活化的巨噬細胞與drg的粘附
結果
1.模型
背柱破碎損傷后,再生的軸突會遭遇巨噬細胞和小神經膠質并形成營養不良的末端。這一點示意性地示于圖1。發明人實驗室以前的工作已經表明,巨噬細胞的滲入與背柱破碎損傷后的軸突頂梢枯死相關(圖2和3)。在表征損傷后上行脊柱感覺軸突的軸突頂梢枯死的程度后,本發明人建立了頂梢枯死的體外模型,其可用于評估多種治療方法。所述體外測定法包括在如下基底上培養的成體drg神經元,所述基底具有生長促進蛋白層粘連蛋白和強抑制性硫酸軟骨素蛋白聚糖聚集蛋白聚糖的相反梯度(tometal.,2004)。此斑點梯度足以使軸突生長停止并誘導與在受損傷的脊髓中出現的生長錐類似的營養不良的生長錐的形成。
縮時顯微術使得發明人能夠密切地檢查生長錐動力學,例如絲狀偽足的數目、板層形偽足的程度和營養不良末端中囊泡的數目。在營養不良的軸突與巨噬細胞之間經常出現導致軸突大幅回縮的直接的細胞-細胞接觸(圖4和5)。直接的細胞接觸是誘導回縮所需的,因為巨噬細胞條件培養基和營養不良軸突附近巨噬細胞的存在均未導致回縮。因此,發明人假設活化的巨噬細胞的清除或調制可成為脊髓損傷中可能的治療靶標。
發明人已經闡明了巨噬細胞-神經元相互作用導致頂梢枯死的機制。已知巨噬細胞分泌多種蛋白酶,這些蛋白酶輔助碎片的分解和清除,并且發明人已證明巨噬細胞會表達和分泌mmp-9。發明人假設某種蛋白酶能夠負責將營養不良的軸突局部地移離基底導致其回縮。巨噬細胞表達的一類蛋白酶是基質金屬蛋白酶(mmp)。已知mmp是導致cns再生障礙的原因,因為缺少某些mmp的轉基因小鼠在損傷后表現出強的軸突再生長,如用通用mmp抑制劑gm6001處理的動物的表現一樣。在加入巨噬細胞時,將gm6001——其在mmp活性位點作為鋅螯合劑——施加于縮時培養皿中。在體外模型中用gm6001或特異性mmp-9抑制劑處理(圖6)阻止了與巨噬細胞直接的細胞-細胞接觸后營養不良的生長錐的回縮,而特異性mmp-2抑制劑沒有阻止。gm6001或所述特異性mmp-9抑制劑不阻止巨噬細胞與營養不良的軸突之間直接的細胞-細胞接觸。因此,證明mmp和mmp-9在軸突頂梢枯死中起作用。
2.mapc在體外阻止巨噬細胞介導的軸突頂梢枯死
圖7示出了為確定mapc是否能調制巨噬細胞的抑制性作用的實驗設計的示意圖。將mapc加至1divdrg斑點培養物并再孵育1天。這些共培養神經元的生長錐的形態迥異于通常出現于斑點上的營養不良的生長錐。這些生長錐的能動性增加、變平并具有大量的板層形偽足。巨噬細胞接觸所述生長錐和軸突,但這些接觸經常是短暫的,并且成像的6個軸突中有5個不發生特征性巨噬細胞介導的回縮(圖8)。
此結果可能是由于mapc的神經刺激作用或免疫調制作用,或是由于兩種作用的結合。為解決此問題,進行了一系列條件培養基實驗。用mapc條件培養基將解離的drg神經元處理24小時并與培養基對照進行比較以評定最長的神經突。在組之間未觀察到神經突長度的顯著差異,表明此作用可能不完全是神經刺激作用。
直接向所述縮時培養皿中加入mapc條件培養基導致生長錐形態的改變,從營養不良的、停滯的狀態變為能動的、扁平的狀態。巨噬細胞仍然接觸這些軸突,但接觸通常是瞬時的并且通常不導致軸突回縮。以mapc條件培養基預處理的巨噬細胞也在所述斑點上接觸軸突,但不引起回縮(圖9-12)。mapc可能作用于巨噬細胞從而改變它們的受體表達、對損傷細胞的響應或者mmp-9的分泌。
3.mapc降低背柱破碎損傷后軸突頂梢枯死的程度
使用脊髓損傷的背柱破碎模型研究了體內mapc對軸突頂梢枯死的免疫調制作用。軸突頂梢枯死的最劇烈階段發生在損傷后2天至4天之間,這在時空上與活化的巨噬細胞滲入損傷一致。mapc可能會以降低軸突頂梢枯死量的方式調制損傷內活化的巨噬細胞。因此,在損傷后立即將mapc移植到脊髓內并在損傷后2天和4天測量軸突頂梢枯死的程度。將mapc移植到位于損傷尾側大約500微米和中線側面500微米的位置。選擇此位置是為了將mapc置于接近受損傷軸突的末端,以使上行通道的進一步破壞最小化,并且以防止細胞在損傷位置被血液和csf流直接從脊髓中替換。
所移植的mapc成功地整合到脊髓組織中,這一點被在損傷后2天和4天在注射位點處均存在gfp+細胞所證明。此外,mapc從移植位點大幅移出并且能夠占據損傷中心并且還被觀察到與受損傷軸突的末端相關連。在損傷后2天,經mapc移植的動物中軸突頂梢枯死程度與對照動物的軸突頂梢枯死程度無顯著差別(圖13)。mapc移植不阻止在損傷后2天正常出現的軸突頂梢枯死程度。然而,此頂梢枯死的初始階段最可能是由于固有神經元機制,而不受活化的巨噬細胞的介導,因為此時巨噬細胞尚未滲入損傷。
在損傷后4天,與未注射的對照相比,經mapc移植的動物顯示出軸突頂梢枯死程度的明顯下降(圖13)。mapc的移植幾乎完全消除了在此時正常觀察到的軸突頂梢枯死,此研究已經表明軸突頂梢枯死是由活化的巨噬細胞的滲入直接導致的。因此,mapc在受損傷的脊髓內的存在足以減少體內巨噬細胞誘導的軸突頂梢枯死。
實施例iv
損傷中心的波形蛋白/ng2+少突膠質細胞前體細胞在巨噬細胞滲入的時間前后開始擴增,并且神經軸索斷裂的纖維的末端與此細胞群體相關。這表明cns損傷內的ng2+細胞起穩定軸突作用,這使它們成為阻止巨噬細胞介導的回縮的理想候選者。在體外培養1天后向drg培養物加入來自成年小鼠脊髓的ng2+神經膠質細胞。在第2天,在30分鐘的基線觀察期后,將nr8383巨噬細胞加入縮時培養皿中并再觀察2.5小時。與drg共培養的ng2+神經膠質細胞的存在不足以阻止巨噬細胞誘導的軸突回縮(n=5)。在圖14中,在巨噬細胞接觸后軸突回縮并在ng2+膠質細胞上穩固。
方法
1.drg解離
如現有技術所述收集drg(tometal.,2004;daviesetal.,1999)。簡言之,從成年雌性sprague-dawley大鼠(harlan)上切下drg。除去中心根和外周根,并在膠原酶ii(200u/ml,worthington)和dispaseii(2.5u/ml,roche)的hbss溶液中孵育神經節。將消化的drg沖洗并在新鮮hbss-cmf中輕輕研磨三次,然后低速離心。然后將解離的drg重懸浮在補充有b-27、glutamax和青霉素/鏈霉素(均來自invitrogen)的neurobasal-a培養基中并計數。將drg以3000個細胞/ml的密度鋪板于delta-t培養皿上(fisher,pittsburgh,pa),共6000個細胞/培養皿。
2.縮時培養皿制備
與tometal.,2004所述類似地制備delta-t細胞培養皿(fisher)。簡言之,用2號鉆頭通過每個培養皿的上半部鉆一個洞,以形成在縮時顯微術過程中向培養物中添加細胞的進口。然后,將培養皿用無菌水沖洗并在室溫下用聚-l-賴氨酸(0.1mg/ml,invitrogen)包被過夜。然后用無菌水沖洗培養皿并使其干燥。通過吸取2.0ul的集聚蛋白聚糖溶液(2.0mg/ml,sigma,溶于hbss-cmf,invitrogen)于培養物表面上并使其干燥,形成集聚蛋白聚糖梯度斑點。每個培養皿中放置6個斑點。在集聚蛋白聚糖斑點完全干燥后,將培養皿的整個表面在層粘連蛋白溶液(10ug/ml,bti的hbss-cmf溶液)中于37℃溫浴3小時。然后撤除層粘連蛋白浴,立即將細胞鋪板。
3.細胞系巨噬細胞培養物
如yinetal.,2003中所述培養成體sprague-dawley肺泡巨噬細胞系nr8383細胞(atcc#crl-2192)。簡言之,在未包被的組織培養燒瓶(corning)中,將細胞在補充有15%fbs(sigma)、glutamax、penn/strep(invitrogen)和碳酸氫鈉(sigma)的f-12k培養基(invitrogen)中培養,并每周加料2-3次。為制備用于縮時顯微術實驗的細胞系巨噬細胞,用胰蛋白酶/edta(invitrogen)收集細胞,洗滌3次,然后以1.0×106/ml的密度鋪板于含無血清f-12k的未包被組織培養燒瓶中。在用于縮時實驗前,用edta和細胞刮刀收集培養的細胞系巨噬細胞并以2.5×105/70ul的密度重懸浮于添加有hepes(50um,sigma)的neurobasal-a中。
4.mapc培養物
在大鼠mapc培養基中培養被gfp標記的sprague-dawley大鼠mapc,所述培養基由低葡萄糖dmem(invitrogen)、0.4×mcdb-201培養基(sigma)、1×its液體培養基添加物(sigma)、1mg/ml亞油酸-白蛋白(sigma)、100u/ml青霉素g鈉鹽或100μg/ml硫酸鏈霉素(invitrogen)、100μm2-p-l-抗壞血酸(sigma)、100ng/mlegf(sigma)、100ng/mlpdgf(r&dsystems)、50nm地塞米松(sigma)、1000u/mlesgro(chemicon)和2%胎牛血清(hyclone)組成。以1000個細胞/cm2的初始密度將所述培養物鋪板于10ng/ml纖連蛋白(invitrogen)包被的150cm2組織培養燒瓶(corning)中,隨后以200個細胞/cm2重新鋪板。將細胞在37℃和5.0%co2維持于15ml的培養基/燒瓶中,使用胰蛋白酶/edta(invitrogen)每3-4天進行傳代。
5.mapc條件培養基
如上述培養細胞,并在48小時后將條件培養基收集到50ml錐形管(bdbioscience)中。將所述條件培養基在4℃以400×g旋轉5分鐘,然后將上清液移至新的50ml錐形管中。然后將此條件培養基保存于4℃。
如上所述獲得了mapc條件培養基,并將其用amiconmicroconultracelym-33,000mwco離心過濾器濃縮50倍。
6.mapc條件培養基處理的巨噬細胞
如上述培養nr8383大鼠巨噬細胞。在縮時顯微術實驗前的1天,用胰蛋白酶/edta(invitrogen)收集巨噬細胞,洗滌3次,然后以1.0×106/ml的密度鋪板于含無血清f-12k的未包被組織培養燒瓶中。每1ml無血清f12k培養基中加入20ul的50倍所述濃縮mapc條件培養基,終濃度為1×。在用于縮時實驗前,用edta和細胞刮刀收集培養的細胞系巨噬細胞并以2.5×105/70ul的密度重懸浮于添加有hepes(50um,sigma)的neurobasal-a中。
7.縮時顯微術
將drg神經元于37℃孵育48小時,然后進行縮時成像。將含有hepes(50um,sigma)的neurobasal-a培養基加至培養物中,然后轉至加熱的臺架儀器中。使用zeissaxiovert405m顯微鏡和100×油浸物鏡,每30秒取得縮時成像,持續3小時。選擇徑直延伸到斑點邊緣并具有特征營養不良形態的生長錐,觀察神經元30分鐘以確定基線表現,然后加入細胞或條件培養基,然后觀察3小時。
對于細胞加入實驗,從組織培養燒瓶中收集所培養的大鼠源mapc,洗滌3次并重懸浮于neurobasal-a培養基中。對于共培養實驗,在24小時后將mapc(100,000個/培養皿)加至背根神經節神經元培養物中并于37℃再孵育24小時,然后進行縮時成像。
對于在縮時成像過程中將mapc條件培養基加至drg培養物中的實驗,在觀察基線生長錐30分鐘后加入90ul的50×mapc-cm。
在30分鐘基線成像后,將mapc條件培養基處理的巨噬細胞加至縮時培養物中(500,000個細胞/培養皿)。
使用metamorph軟件分析延伸/回縮、生長速率、彎向和分枝。
8.免疫化學檢測
在縮時成像后,將drg在4%pfa中固定并用抗iii型b-微管蛋白(1:500;sigma)、抗硫酸軟骨素(cs-56,1:500,sigma)和抗gfp(1:500,invitrogen)進行免疫染色。
9.原代骨髓源巨噬細胞培養
如tobianetal.,2004中所述收集骨髓祖細胞。簡言之,將股骨從成年雌性sprague-dawley大鼠(harlan)取下。除去股骨末端,將含有補充有10%fbs、glutamax、penn/strep、β-巰基乙醇和hepes(invitrogen)(d10f)的冷dmem的注射器插入股骨,沖出骨髓并收集。然后使所得細胞混合物通過70微米的濾器并離心。然后除去上清液,將所得細胞片狀沉淀物重懸浮于akt裂解緩沖液(biowhitacre)中以裂解紅細胞,然后離心。除去上清液并將含有骨髓前體細胞的片狀沉淀物重懸浮并鋪板于另補充有20%ladmac細胞系條件培養基(cliffordharding贈予)的上述dmem中以誘導分化為巨噬細胞。在培養第10天收集細胞進行實驗。縮時實驗前的1天,用胰蛋白酶/edta收集原代巨噬細胞,用d10f洗滌3次并以1.0×106/ml的密度鋪板于含d10f的未包被陪替氏培養皿(falcon)中。第2天,用edta和細胞刮刀收集原代巨噬細胞并以5.0×105/70ul的密度重懸浮于加hepes的neurobasal-a中,用于縮時顯微術實驗。
10.背柱破碎損傷模型
對于所有手術步驟,均用吸入的異氟醚氣(2%)麻醉成年雌性sprague-dawley大鼠(250-300g)。進行t1椎板切除術來暴露c8脊髓節的背面。用30號針頭在距離中線0.75mm的兩側進行硬腦膜切開術。然后通過將dumont#3jeweler鑷子在c8處插入背脊至1.0mm的深度;壓住鑷子,保持壓力10秒鐘并再重復2次來制造背柱破碎損傷。通過觀察到白色物質的清除來確認損傷的完成。然后用凝膠膜覆蓋硬腦膜上的洞。用4-0尼龍線縫合肌肉層,然后用外科封縫釘閉合皮膚。在閉合切口后,動物沿切口皮下接受麻卡因(1.0mg/kg)并肌內注射丁丙諾啡(0.1mg/kg)。手術后,在麻醉恢復期間用加熱燈使動物保暖,并使其自由取食和飲水。將動物在損傷后2、4、7、14或28天處死。
11.細胞移植
從組織培養燒瓶中收集培養的大鼠源mapc或原代骨髓源巨噬細胞(已經用干擾素γ和lpc刺激24小時),在hbss-cmf中洗滌3次并以200,000細胞/ul的密度重懸浮于hbss-cmf中。背柱破碎損傷后,立即將1.0ul細胞懸浮液在單側以0.5mm深度注射到右側脊柱中。注射位置在中線側面0.5mm和損傷邊緣尾端0.5mm。通過連接于nanojectii(drummond)的拉制玻璃吸液管以15秒間隔用44次23.0nl的脈沖注射細胞。在最后一次注射后的2分鐘從脊髓中抽出玻璃吸液管。在移植后,用凝膠膜覆蓋注射位點,用4-0ethicon縫線縫合肌肉層,然后用外科封縫釘閉合皮膚。手術后,在麻醉恢復期間用加熱燈使動物保暖,并使其自由取食和飲水。在損傷后2天或4天處死動物。
12.軸突標記
處死前2天,用texas-red綴合的3000mw葡聚糖單側標記脊柱。簡言之,暴露右后肢的坐骨神經,用dumont#3鑷子擠壓3次,持續10秒鐘。在擠壓位點處用hamilton注射器將1.0ul3000mw葡聚糖-texas-red的10%無菌水溶液注射到坐骨神經中。用4-0尼龍線縫合肌肉層,然后用外科封縫釘閉合皮膚。在閉合切口后,動物沿切口皮下接受麻卡因(1.0mg/kg)并肌內注射丁丙諾啡(0.1mg/kg)。手術后,在麻醉恢復期間用加熱燈使動物保暖,并使其自由取食和飲水。將動物在標記后2天用過量異氟醚處死并用pbs繼而用4%pfa灌注。收集組織并在4%pfa中后固定并進行免疫組織化學測定。
13.免疫組織化學測定
將組織在4%pfa后固定過夜,然后浸沒在30%蔗糖中過夜,在oct封裝介質中冷凍,然后在恒切片機上切成20um的縱向切片。然后將組織用抗gfap/alexafluor-405、抗ed-l/alexafluor-594或-633、抗gfp/alexafluor-488和抗波形蛋白/alexafluor-633染色。然后在zeissaxiovert510激光掃描共焦顯微鏡上在10×放大率下成像。
14.軸突頂梢枯死定量
為定量軸突頂梢枯死,對每只動物分析了始于脊髓背面下200um深處的三個連續切片。通過特征性gfap和/或波形蛋白染色模式確定損傷中心,然后使用zeisslsm5imagebrowser軟件確定中心位置。然后測量最遠深入到損傷中的5個標記軸突的末端與損傷中心的距離。將一組中所有動物的所有切片的測量值平均以得出每個時間點的頂梢枯死平均距離。
實施例v
間充質干細胞可商業購得。例如,ratmesenchymalstemcellkit(milliporecatalogno.scr026)提供從成年fisher344大鼠的骨髓分離的即用間充質原代干細胞,以及用于鑒定間充質干細胞群體的一組陽性和陰性選擇標志物。陽性細胞標志物包括針對間充質干細胞上存在的兩種細胞表面分子(整合素b1和cd45)的抗體。陰性細胞標志物包括針對間充質干細胞不表達的兩種特定造血細胞表面標志物(白細胞上存在的cd14以及單核細胞和巨噬細胞上存在的cd45)的抗體。對這些間充質干細胞評定減少回縮的能力,并發現這些間充質干細胞在體外(神經膠質疤痕)減少回縮(減少附著)。
實施例vi
對在5μg/ml層粘連蛋白上培養的成體drg進行mapc條件培養基處理可促進神經突生長。參見圖16。對向其中添加有含有neurobasal-a的培養基并添加有mapc條件培養基、對照培養基或沒有另外添加培養基的各組測量每個解離的drg的最長軸突。所有條件彼此均顯著不同,單因素anova,*p<0.0001。b,表示未處理的drg神經元的平均向外生長量的16×圖像。c,表示用mapc條件培養基預處理的drg的平均向外生長量的16×圖像。
此結果可能是由于mapc的神經刺激作用或免疫調制作用,或者是由于兩種作用的結合。為解決此問題,進行了一系列條件培養基實驗。用新鮮mapc條件培養基將解離的drg神經元處理24小時并與培養基對照進行比較。測量每個drg的最長神經突。新鮮mapc條件培養基顯著增加了層粘連蛋白上的向外生長。這表明mapc分泌一種或多種對成體神經元具有神經刺激作用的生長因子。預先冷凍的mapc條件培養基對神經元不具有同樣的作用,表明所述因子在冷凍過程中改變或失活。
參考文獻
arantesetal.,neuroreport11:177-181(2000)
bakeretal.,expneurol205:82-91(2007)
bakerandhagg,jneurotrauma22:415-428(2005)
baschetal.,(1983)
batinic,d.,etal.,(1990)
borgensetal.,jcompneurol250:157-167(1986)
brownetal.,eurjneurosci9:2057-2063(1997)
buschandsilver,curropinneurobiol17:120-127(2007)
caiz.h.,etal.,(1988)
caoetal.,neurorehabilneuralrepair(2007)
chang,p.,etal.,(1999)
chang,t.m.,(1992)
daviesetal.,jneurosci19:5810-5822(1999)
deetal.,jneuropatholexpneurol61:237-244(2002)
desimoneetal.,molneurobio29:197-212(2004)
donnellyandpopovich,expneurol209:378-388(2008)
farretal.,aplysiajneurobiol46:89-96(2001)
fayazandtator,jneuroscimethods102:69-79(2000)
fishmanandkelley,neurology34:1161-1167(1984)
fitchandsilverexpneurol148:587-603(1997)
fitchetal.,jneurosci19:8182-8198(1999)
grimpeandsilver,jneurosci24:1393-1397(2004)
hilletal.,expneurol171:153-169(2001)
hohlfeldetal.,neurology68:s58-s63(2007)
houleandjin,expneurol169:208-217(2001)
iizukaetal.,jneurosurg66:595-603(1987)
jonesetal.,expneurol182:399-411(2003)
kelmetal.,currbiol4:965-972(1994)
kerschensteineretal.,natmed11:572-577(2005)
kwonetal.,procnatlacadsciusa99:3246-3251(2002)
liandraisman,expneurol134:102-111(1995)
liuetal.,jneurosci26:3087-3101(2006)
longbrakeetal.,jneurochem102:1083-1094(2007)
matthew,h.w.,etal.,(1991)
mckeonetal.,jneurosci11:3398-3411(1991)
mcphailetal.,eurj.neurosci20:1984-1994(2004)
morgensternetal.,progbrainres137:313-322(2002)
muschler,g.f.,etal.,(1997)
oudegaetal.,eurj.neurosci11:2453-2464(1999)
prewittetal.,expneurol148:433-443(1997)
popovichetal.,jcompneurol377:443-464(1997)
popovichandlongbrake,natrevneurosci9:481-493(2008)
popovichetal.,expneurol158:351-365(1999)
raffetal.,science296:868-871(2002)
ramonycajal,london:oxfordup(1928)
rapalinoetal.,natmed4:814-821(1998)
schwartzetal.,neurosurgery44:1041-1045(1999b)
schwartzetal.,amjneuroradial26:7-18(2005)
seifctal.,jneurotrauma24:1513-1528(2007)
silverandmiller,natrevneurosci5:146-156(2004)
snowetal.,expneurol109:111-130(1990)
sobeletal.,expneurol54:202-213(1995)
steinmetzetal.,jneurosci25:8066-8076(2005)
stirlingetal,jneurosci24:2182-2190(2004)
tangetal.,jneuroscires71;427-444(2003)
tangetal.,jcellbiol138:1355-1366(1997)
tobianetal.,jimmunol172:5277-5286(2004)
tometal.,jneurosci24:6531-6539(2004)
umeharaetal.,immunolcellbiol79:298-302(2001)
vanrooijenetal.,trendsbiotechnol15:178-185(1997)
vanrooijenandvankesteren-hendrikx,jliposomeres12:81-94(2002)
vanrooijenandsanders,jimmunomethods174:83-93(1994)
watanabeetal.,jneurotrauma16:255-265(1999)
wysockiandsato,(1978)
xieandzheng,expneurol209:302-312(2008)
yanagi,k.etal.,(1989)
yinetal.,j.neurosci23:2284-2293(2003)
yiuandhe,natrevneurosci7:617-627(2006)
zujovicetal.,glia29:305-315(2000)
- 還沒有人留言評論。精彩留言會獲得點贊!
- SPNP?IX在制備Kv4.2通道工具試劑中的應用的制造方法與工藝
- SPNP?IX在制備Kv4.3通道工具試劑中的應用的制造方法與工藝
- SPNP?27在制備Nav1.5通道抑制劑中的應用的制造方法與工藝
- SPNP?27在制備Nav1.3通道抑制劑中的應用的制造方法與工藝
- 一種多肽在制備Nav1.3鈉通道工具試劑中的應用的制造方法與工藝
- SPKP?XI在制備Kv4.2通道抑制劑中的應用的制造方法與工藝
- 基于水凝膠溶脹特性的可控自變形神經微電極的制造方法與工藝
- SPKP?XI在制備Kv4.1鉀通道抑制劑中的應用的制造方法與工藝
- 小駁骨提取物作為神經保護劑的應用的制造方法與工藝
- 巴戟天低聚果糖提取物在制備防治阿爾茨海默癥藥物、保健品或食品中的應用的制造方法與工藝