GhPRE1基因及其在改良棉纖維品質中的應用的制作方法
本發明涉及植物學領域,更具體地涉及ghpre1基因及其改良棉纖維品質中的應用。
背景技術:
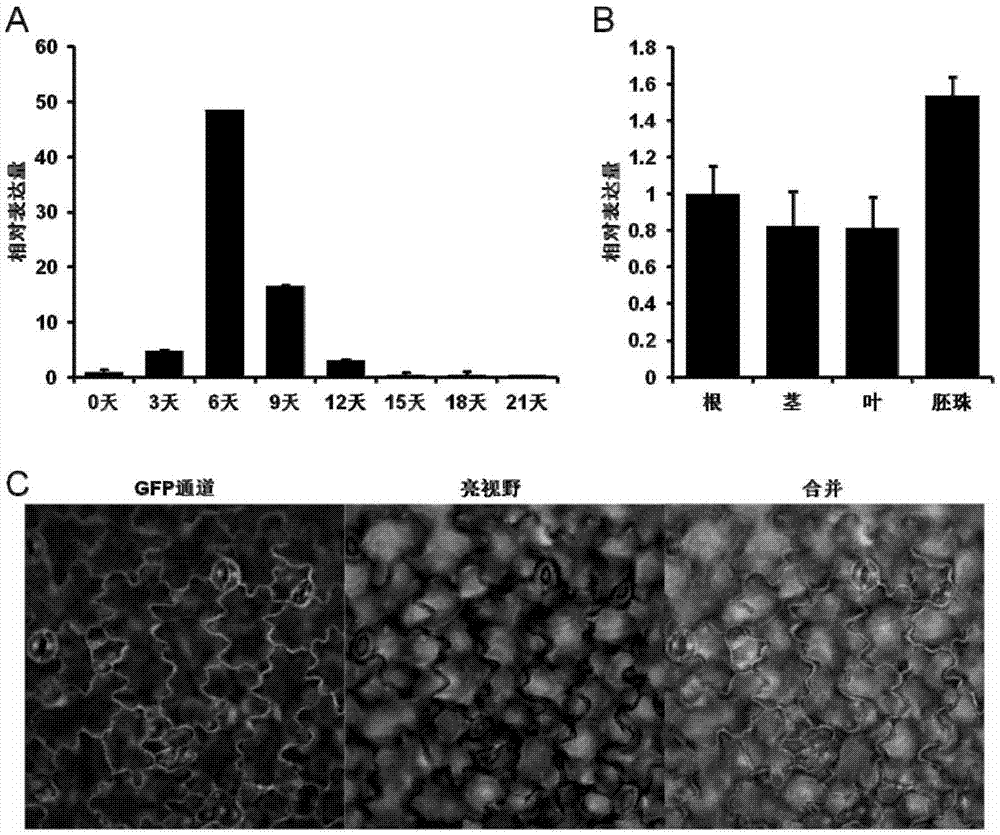
:棉花是世界上最主要的農作物之一,產量大、生產成本低,使棉制品價格比較低廉。棉纖維能制成多種規格的織物,從輕盈透明的巴里紗到厚實的帆布和厚平絨,適于制作各類衣服、家具布和工業用布。棉織物堅牢耐磨,能夠洗滌和在高溫下熨燙,棉布由于吸濕和脫濕快速而使穿著舒服,應用廣泛。我國是世界上棉花產量最高的國家之一。棉纖維是由棉花種子表皮細胞分化產生的,為極度伸長的單細胞結構。其分化和發育過程可分為:纖維細胞分化與突起、纖維細胞的迅速伸長、次生壁的合成和脫水成熟等4個時期。纖維細胞各個時期的生命活動影響棉纖維的產量和品質形成,其中纖維伸長和次生壁合成兩個時期對纖維的發育與品質形成關系最為密切。棉纖維發育和結構的特點是由一系列在特定的時間和空間表達的基因決定的,而基因表達的時空特異性主要是由轉錄調控因子與下游基因的啟動子之間的相互作用實現的。克隆鑒定棉纖維發育的關鍵調控基因,并對其功能進行研究,認識棉纖維發育的分子調控機理,對于改善棉纖維品質、提高作物產量等具有十分重要的生物學意義和應用價值。因此,本領域迫切需要開發新的調控棉纖維發育的基因,并對其功能應用進行相應的研究。技術實現要素:本發明的目的在于提供一種調控棉纖維發育的ghpre1基因及其在改良棉纖維品質中的應用。在本發明的第一方面,提供了一種ghpre1基因或其編碼蛋白的用途,所述的ghpre1基因或其編碼蛋白用于選自下組的用途:(a)用于制備促進棉屬植物棉纖維伸長的試劑或組合物;(b)用于改善棉屬植物的棉纖維長度性狀;(c)用于增加棉屬植物棉纖維的長度;(d)用于增強棉屬植物棉纖維的斷裂比強度;(e)用于改善棉屬植物棉纖維的馬克隆值;(f)用于提高棉屬植物棉纖維的紡紗均勻性。在另一優選例中,所述的ghpre1基因或其編碼蛋白還用于選自下組的一種或多種用途:(d)用作制備促進植物細胞伸長的試劑或組合物;和(e)用于增加植物細胞的長度。在另一優選例中,所述的植物為棉屬植物。在另一優選例中,所述的“改善棉屬植物的棉纖維長度性狀”或所述的“增加棉屬植物棉纖維的長度”指:導入外源的所述“ghpre1基因”所獲得的轉基因棉屬植物的棉纖維長度l1與未導入外源的所述“ghpre1基因”的同一種類的棉屬植物的棉纖維長度l0之比值(l1/l0)為105%-120%,較佳地為110%-112%。在另一優選例中,所述的“增強棉屬植物棉纖維的斷裂比強度”指:導入外源的所述“ghpre1基因”所獲得的轉基因棉屬植物的棉纖維斷裂比強度s1與未導入外源的所述“ghpre1基因”的同一種類的棉屬植物的棉纖維斷裂比強度s0之比值(s1/s0)為103%-120%,較佳地為105%-112%。在另一優選例中,所述的“增強棉屬植物棉纖維的紡紗均勻性”指:導入外源的所述“ghpre1基因”所獲得的轉基因棉屬植物的棉纖維紡紗均勻性a1與未導入外源的所述“ghpre1基因”的同一種類的棉屬植物的棉纖維紡紗均勻性a0之比值(a1/a0)為105%-125%,較佳地為110%-115%。在另一優選例中,所述的“改善棉屬植物棉纖維的馬克隆值”指導入外源的所述“ghpre1基因”所獲得的轉基因棉屬植物的棉纖維馬克隆值處于國家b2級標準。在另一優選例中,所述的ghpre1基因包括野生型ghpre1基因和突變型ghpre1基因。在另一優選例中,所述的突變型包括突變后編碼蛋白的功能未發生改變的突變形式(即功能與野生型編碼蛋白相同或基本相同)。在另一優選例中,所述的突變型ghpre1基因編碼的多肽與野生型ghpre1基因所編碼的多肽相同或基本相同。在另一優選例中,所述的突變型ghpre1基因包括與野生型ghpre1基因相比,同源性≥80%(較佳地≥90%,更佳地≥95%)的多核苷酸。在另一優選例中,所述的突變型ghpre1基因包括在野生型ghpre1基因的5'端和/或3'端截短或添加1-60個(較佳地1-30,更佳地1-10個)核苷酸的多核苷酸。在另一優選例中,所述的基因包括dna、cdna、和/或mrna。在另一優選例中,所述的ghpre1基因的cds序列如seqidno.:1所示。在另一優選例中,所述的ghpre1基因的編碼蛋白如seqidno.:2所示。在另一優選例中,所述的ghpre1基因的基因組序列如seqidno.:3所示。在另一優選例中,所述的ghpre1基因來源于棉屬植物。在另一優選例中,所述的棉屬植物為棉花。在另一優選例中,所述的包括陸地棉、海島棉、亞洲棉和草棉。在另一優選例中,所述的棉纖維為棉屬(gossypium)植物的種籽纖維。在另一優選例中,所述的ghpre1基因具有促進棉纖維伸長的功能。在另一優選例中,所述的細胞選自下組:纖維細胞、胚軸細胞、根細胞、種子細胞、和葉柄細胞。在本發明的第二方面,提供了一種調控棉屬植物的棉纖維長度性狀的方法,包括步驟:(a)將外源的構建物導入植物細胞,其中所述的構建物含有外源的ghpre1基因序列、促進ghpre1基因表達的外源核苷酸序列、或抑制ghpre1基因表達的外源核苷酸序列,從而獲得導入外源構建物的植物細胞;(b)將上一步驟獲得的所述導入外源構建物的植物細胞,再生成植株:和(c)任選地對所述再生的植株進行性狀鑒定,從而獲得性狀改良的植物。在另一優選例中,所述外源的ghpre1基因序列還包含與orf序列操作性連接的啟動子和/或終止子。在另一優選例中,所述的啟動子選自下組:組成型啟動子、組織特異性啟動子、誘導型啟動子、和強啟動子。在另一優選例中,所述的組成性啟動子包括35s啟動子。在另一優選例中,所述的組織特異性啟動子包括rdl1啟動子。在另一優選例中,所述的終止子選自下組:nos終止子。在另一優選例中,所述的外源核苷酸序列包括干擾所述ghpre1基因表達的核苷酸序列。在另一優選例中,所述的外源核苷酸序列包括rna干擾序列。在本發明的第三方面,提供了一種促進棉屬植物的棉纖維伸長的方法,所述方法包括以下步驟:在所述棉屬植物中,促進ghpre1基因的表達或促進ghpre1蛋白的活性。在另一優選例中,所述的表達為棉纖維組織中的表達。在另一優選例中,所述的表達為棉纖維組織發育期的表達。在另一優選例中,所述的棉纖維組織發育期的表達為棉纖維發育第1-30天,較佳地2-20天,更佳地3-12天的表達。在另一優選例中,所述方法包括給予植物ghpre1基因或其編碼的多肽的促進劑。在另一優選例中,所述方法包括向植物中導入外源的ghpre1基因。在另一優選例中,所述方法包括步驟:(i)提供一植物或植物細胞;和(ii)將ghpre1基因序列導入所述植物或植物細胞,從而獲得轉基因的植物或植物細胞。在另一優選例中,所述方法包括步驟:(a)提供攜帶ghpre1基因序列的表達載體的農桿菌;(b)將植物細胞或組織或器官與步驟(a)中的農桿菌接觸,從而使ghpre1的基因序列轉入植物細胞,并且整合到植物細胞的染色體上;(c)選擇已轉入ghpre1基因序列的植物細胞或組織或器官;和(d)將步驟(c)中的植物細胞或組織或器官再生為植株。在本發明的第四方面,提供了一種促進棉屬植物細胞伸長的方法,所述方法包括以下步驟:促進所述細胞中ghpre1基因的表達或促進ghpre1蛋白的活性。在另一優選例中,所述方法包括給予細胞ghpre1基因或其編碼的多肽的促進劑。在另一優選例中,所述方法包括向細胞中導入外源的ghpre1基因。在本發明的第五方面,提供了一種ghpre1基因或其編碼蛋白的調控劑的用途,用于選自下組的用途:(a)用于調控棉屬植物的棉纖維長度性狀,或用于制備調控棉纖維長度的試劑或組合物;(b)用于調控棉屬植物的棉纖維的斷裂比強度,或用于制備調控棉纖維斷裂比強度的試劑或組合物;(c)用于調控棉屬植物的棉纖維的馬克隆值,或用于制備調控棉纖維馬克隆值的試劑或組合物;(d)用于調控棉屬植物的棉纖維的紡紗均勻性,或用于制備調控棉纖維紡紗均勻性的試劑或組合物。在另一優選例中,所述的組合物包括農用組合物。在另一優選例中,所述的調控劑包括促進劑、抑制劑。在另一優選例中,所述的調控劑為促進劑,并且所述的調控是指促進所述棉纖維伸長。在另一優選例中,所述的調控劑為抑制劑,并且所述的調控是指抑制所述棉纖維伸長。在另一優選例中,所述的調控劑包括小分子化合物、或核酸類物質。在另一優選例中,所述的核酸類物質選自下組:mirna、shrna、sirna、或其組合。在本發明的第六方面,提供了一種增強棉屬植物棉纖維的斷裂比強度的方法,所述方法包括以下步驟:在所述棉屬植物中,促進ghpre1基因的表達或促進ghpre1蛋白的活性。在本發明的第七方面,提供了一種改善棉屬植物棉纖維的馬克隆值的方法,所述方法包括以下步驟:在所述棉屬植物中,促進ghpre1基因的表達或促進ghpre1蛋白的活性。在本發明的第八方面,提供了一種提高棉屬植物棉纖維的紡紗均勻性的方法,所述方法包括以下步驟:在所述棉屬植物中,促進ghpre1基因的表達或促進ghpre1蛋白的活性。應理解,在本發明范圍內中,本發明的上述各技術特征和在下文(如實施例)中具體描述的各技術特征之間都可以互相組合,從而構成新的或優選的技術方案。限于篇幅,在此不再一一累述。附圖說明圖1顯示了轉基因載體35s::ghpre1、35s::dsghpre1、rdl1::ghpre1的構建。其中,圖1a顯示了轉基因所用商業化載體pcambia2301空載體圖譜;圖1b顯示了轉基因所用商業化載體pbi121空載體圖譜;1c顯示了載體35s::ghpre1、rdl1::ghpre1、35s::dsghpre1、35s::ghpre1-venus的構建示意圖。圖2顯示了ghpre1基因表達特征和蛋白亞定位。其中,圖2a顯示了ghpre1基因在棉纖維不同發育時期的表達特征;圖2b顯示了ghpre1基因在棉花不同組織中的表達量;圖2c顯示了c末端融合gfp的ghpre1-gfp蛋白在煙草表皮細胞中的亞細胞定位。圖3顯示了35s::ghpre1轉基因擬南芥表型分析。其中,圖3a顯示了 35s::ghpre1轉基因植物萌發后5天的小苗表現出下胚軸和根等部位顯著伸長的特征,l2、l15和l16為三個獨立的轉基因株系;圖3b顯示了35s::ghpre1轉基因植物中ghpre1表達量的檢測;圖3c顯示了生長3周的35s::ghpre1和35s::ghpre1-venus轉基因擬南芥葉柄表現出增長的表型。圖4顯示了rdl1::ghpre1,35s::dsghpre1轉基因棉花的pcr分子鑒定。其中,圖4a顯示了rdl1::ghpre1轉基因棉花陽性鑒定結果;圖4b顯示了35s::dsghpre1轉基因棉花陽性鑒定結果。其中的pcr鑒定采用nptii特異的引物,載體泳道(正對照)為以pcambia2301載體為模板擴增的產物。圖5顯示了rdl1::ghpre1轉基因棉花的纖維長度分析和qrt-pcr分子鑒定。其中,圖5a顯示了rdl1::ghpre1轉基因棉花表現出成熟棉纖維增長的表型;圖5b顯示了rdl1::ghpre1轉基因棉花纖維長度統計結果;圖5c顯示了qrt-pcr檢測rdl1::ghpre1轉基因棉花開花后6天纖維細胞中ghpre1的表達量。圖6顯示了35s::dsghpre1轉基因棉花的纖維長度分析和qrt-pcr分子鑒定。其中,圖6a顯示了35s::dsghpre1轉基因棉花表現出成熟棉纖維增長的表型;圖6b顯示了35s::dsghpre1轉基因棉花纖維長度統計結果;圖6c顯示了qrt-pcr檢測35s::dsghpre1轉基因棉花開花后6天纖維細胞中ghpre1的表達量。圖7顯示了ghacy轉基因棉花的分子鑒定和纖維表型分析。其中圖a顯示了ghacy基因在開花后3-12天的纖維細胞中高表達;圖b顯示了35s::dsghacy轉基因棉花幼葉中ghacy的表達量分析;圖c顯示了35s::dsghacy轉基因棉花纖維長度有輕微變短的表型;圖d顯示了35s::ghacy轉基因棉花中ghacy的表達分析;圖e顯示了35s::ghacy轉基因棉花纖維長度無明顯變化。圖8顯示了大田種植的rdl1::ghpre1轉基因棉花的纖維品質檢測結果。從中可以看出2014年和2015年種植的rdl1::ghpre1轉基因棉花纖維長度、斷裂比強度、馬克隆值和紡紗均勻性等品質穩定地優于對照組的品質,說明利用ghpre1改良的棉花在農業生產和棉紡工業中具有重要的應用前景。具體實施方式本發明人經過廣泛而深入地研究,首次發現了一種能夠調控棉纖維發育的ghpre1基因,以及ghpre1基因在改良棉纖維長度、斷裂比強度、馬克隆值、紡紗均勻性等纖維品質中的應用。實驗表明:采用棉纖維細胞特異的rdl1啟動子增強ghpre1基因的表達,可以改良棉纖維長度性狀,斷裂比強度、馬克隆值和紡紗均勻性等品質,獲得了纖維品質顯著改良的轉基因棉花。在此基礎上完成了本發明。棉花棉花是錦葵科棉屬植物,是一種重要的經濟作物,其種子纖維為市場提供大量天然紡織原料。植株灌木狀,在熱帶地區栽培可長到6米高,一般為1到2米。花朵乳白色,開花后不久轉成深紅色然后凋謝,留下綠色小型的蒴果,稱為棉鈴。棉鈴內有棉籽,棉籽上的茸毛從棉籽表皮長出,塞滿棉鈴內部。棉鈴成熟時裂開,露出柔軟的棉纖維。常見棉纖維白色至白中帶黃,長約2至4厘米,含纖維素約87~90%。陸地棉(gossypiumhirsutum)為四倍體栽培棉種,又名:大陸棉、美洲棉、墨西哥棉等,是世界上四大棉花栽培種中重要的品種,在中國產棉區普遍栽培。葉較大,3-5裂,裂刻長度不及葉片長度的1/2,有苞葉外蜜腺,花冠較大,乳白色,通常無紅心,鈴大,卵圓形,鈴面腺體少,較光滑,種子中等大小,一般為毛籽,纖維長度約為21-32mm,纖維細度為4500-7000m/g,纖維直徑為13.5-19μm,單纖維強力為3.5-4.5g。ghpre1基因如本文所用,術語“ghpre1基因”、“棉纖維長度相關基因”、“本發明基因”可以互換使用,都是指本發明的具有調控棉纖維長度功能的基因。如本文所用,術語“特異性表達”是指目的基因在植物中特定的時間和/或特定的組織的表達。如本文所用,“外源的”或“異源的”是指不同來源的兩條或多條核酸或蛋白質序列之間的關系。例如,如果啟動子與目的基因序列的組合通常不是天然存在的,則啟動子對于該目的基因來說是外源的。特定序列對于其所插入的細胞或生物體來說是“外源的”。本發明利用數字基因表達譜分析技術和rt-pcr分析從陸地棉中篩選獲得一個特異地在棉纖維伸長期高表達的基因ghpre1,并在擬南芥中異源表達ghpre1證明其確實具有促進細胞伸長生長的功能。在陸地棉中,ghpre1特異地在開花后6~9天快速伸長的棉纖維中高表達。上述結果提示,ghpre1可能特化為纖維細胞生長的調控因子,在棉纖維細胞發育早期發揮轉錄調控作用,正向調節棉纖維的伸長生長。為了進一步深入研究ghpre1的生物學功能,為高產優質棉花品種的培育提供分子依據,本發明進行了轉基因功能分析,構建了ghpre1過表達和rna干擾(rnai)的載體進行了棉花轉化,并成功獲取各個載體的多個轉基因株系。對種植于溫室和大田轉基因植物進行分析,發現過表達ghpre1的轉基因植株,表現出纖維長度顯著增加的表型,而抑制ghpre1表達的轉基因植株,表現出纖維長度顯著變短的表型。這一結果說明轉錄因子ghpre1是棉纖維細胞發育的重要調 控因子,在促進棉纖維伸長和纖維品質改良等方面具有巨大潛力和應用價值。本發明的ghpre1基因可以是dna形式或rna形式。dna形式包括cdna、基因組dna或人工合成的dna。基因組dna可以是與seqidno.:3所示的序列相同或者是簡并的變異體。本發明的dna可以是單鏈的或是雙鏈的,dna可以是編碼鏈或非編碼鏈。編碼成熟多肽的編碼區序列可以與seqidno.:1所示的編碼區序列相同或者是簡并的變異體。如本文所用,“簡并的變異體”在本發明中是指編碼具有seqidno.:2的蛋白質,但與seqidno.:1所示的編碼區序列或seqidno.:3所示的基因組序列有差別的核酸序列。編碼seqidno.:2的成熟多肽的多核苷酸包括:只編碼成熟多肽的編碼序列;成熟多肽的編碼序列和各種附加編碼序列;成熟多肽的編碼序列(和任選的附加編碼序列)以及非編碼序列。術語“編碼多肽的多核苷酸”可以是包括編碼此多肽的多核苷酸,也可以是還包括附加編碼和/或非編碼序列的多核苷酸。本發明還涉及上述多核苷酸的變異體,其編碼與本發明有相同的氨基酸序列的多肽或多肽的片段、類似物和衍生物。此多核苷酸的變異體可以是天然發生的等位變異體或非天然發生的變異體。這些核苷酸變異體包括取代變異體、缺失變異體和插入變異體。如本領域所知的,等位變異體是一個多核苷酸的替換形式,它可能是一個或多個核苷酸的取代、缺失或插入,但不會從實質上改變其編碼的多肽的功能。本發明還涉及與上述的序列雜交且兩個序列之間具有至少50%,較佳地至少70%,更佳地至少80%相同性的多核苷酸。本發明特別涉及在嚴格條件下與本發明所述多核苷酸可雜交的多核苷酸。在本發明中,“嚴格條件”是指:(1)在較低離子強度和較高溫度下的雜交和洗脫,如0.2×ssc,0.1%sds,60℃;或(2)雜交時加有變性劑,如50%(v/v)甲酰胺,0.1%小牛血清/0.1%ficoll,42℃等;或(3)僅在兩條序列之間的相同性至少在90%以上,更好是95%以上時才發生雜交。并且,可雜交的多核苷酸編碼的多肽與seqidno.:2所示的成熟多肽有相同的生物學功能和活性。本發明還涉及與上述的序列雜交的核酸片段。如本文所用,“核酸片段”的長度至少含15個核苷酸,較好是至少30個核苷酸,更好是至少50個核苷酸,最好是至少100個核苷酸以上。核酸片段可用于核酸的擴增技術(如pcr)以確定和/或分離編碼棉纖維相關多肽的多聚核苷酸。ghpre1基因編碼的多肽如本文所用,術語“棉纖維長度相關多肽”、“本發明多肽”、“ghpre1基因編碼的多肽”、“ghpre1基因編碼的蛋白”、“ghpre1多肽”可以互換使用,都是指本發明的、來源于棉屬植物的、具有調控棉纖維長度功能的多肽。本發明的多肽可以是重組多肽、天然多肽、合成多肽,優選重組多肽。本發明的多肽可以是天然純化的產物,或是化學合成的產物,或使用重組技術從原核或真核宿主(例如,細菌、酵母、高等植物、昆蟲和哺乳動物細胞)中產生。根據重組生產方案所用的宿主,本發明的多肽可以是糖基化的,或可以是非糖基化的。本發明的多肽還可包括或不包括起始的甲硫氨酸殘基。本發明還包括棉纖維長度相關多肽的片段、衍生物和類似物。如本文所用,術語“片段”、“衍生物”和“類似物”是指基本上保持本發明的天然棉纖維長度相關多肽相同的生物學功能或活性的多肽。本發明的多肽片段、衍生物或類似物可以是(i)有一個或多個保守或非保守性氨基酸殘基(優選保守性氨基酸殘基)被取代的多肽,而這樣的取代的氨基酸殘基可以是也可以不是由遺傳密碼編碼的,或(ii)在一個或多個氨基酸殘基中具有取代基團的多肽,或(iii)成熟多肽與另一個化合物(比如延長多肽半衰期的化合物,例如聚乙二醇)融合所形成的多肽,或(iv)附加的氨基酸序列融合到此多肽序列而形成的多肽(如前導序列或分泌序列或用來純化此多肽的序列或蛋白原序列,或融合蛋白)。根據本文的教導,這些片段、衍生物和類似物屬于本領域熟練技術人員公知的范圍。在本發明中,術語“棉纖維長度相關多肽”指具有棉纖維長度相關多肽活性的seqidno.:2序列的多肽。該術語還包括具有與棉纖維長度相關多肽相同功能的、seqidno.:2序列的變異形式。這些變異形式包括(但并不限于):一個或多個(通常為1-50個,較佳地1-30個,更佳地1-20個,最佳地1-10個)氨基酸的缺失、插入和/或取代,以及在c末端和/或n末端添加一個或數個(通常為20個以內,較佳地為10個以內,更佳地為5個以內)氨基酸。例如,在本領域中,用性能相近或相似的氨基酸進行取代時,通常不會改變蛋白質的功能。又比如,在c末端和/或n末端添加一個或數個氨基酸通常也不會改變蛋白質的功能。該術語還包括棉纖維長度相關多肽的活性片段和活性衍生物。該多肽的變異形式包括:同源序列、保守性變異體、等位變異體、天然突變體、誘導突變體、在高或低的嚴緊度條件下能與棉纖維長度相關多肽的dna雜交的dna所編碼的蛋白、以及利用抗棉纖維長度相關多肽的抗血清獲得的多肽或蛋白。本發明還提供了其他多肽,如包含棉纖維長度相關多肽或其片段的融合蛋白。除了幾乎全長的多肽外,本發明還包括了棉纖維長度相關多肽的可溶性片段。通常,該片段具有棉纖維長度相關多肽序列的至少約10個連續氨基酸,通常至少約30個連續氨基酸,較佳地至少約50個連續氨基酸,更佳地至少約80 個連續氨基酸,最佳地至少約100個連續氨基酸。本發明還提供棉纖維長度相關多肽或多肽的類似物。這些類似物與天然棉纖維長度相關多肽的差別可以是氨基酸序列上的差異,也可以是不影響序列的修飾形式上的差異,或者兼而有之。這些多肽包括天然或誘導的遺傳變異體。誘導變異體可以通過各種技術得到,如通過輻射或暴露于誘變劑而產生隨機誘變,還可通過定點誘變法或其他已知分子生物學的技術。類似物還包括具有不同于天然l-氨基酸的殘基(如d-氨基酸)的類似物,以及具有非天然存在的或合成的氨基酸(如β、γ-氨基酸)的類似物。應理解,本發明的多肽并不限于上述例舉的代表性的多肽。修飾(通常不改變一級結構)形式包括:體內或體外的多肽的化學衍生形式如乙酰化或羧基化。修飾還包括糖基化。修飾形式還包括具有磷酸化氨基酸殘基(如磷酸酪氨酸,磷酸絲氨酸,磷酸蘇氨酸)的序列。還包括被修飾從而提高了其抗蛋白水解性能或優化了溶解性能的多肽。在本發明中,“棉纖維長度相關多肽保守性變異多肽”指與seqidno.:2的氨基酸序列相比,有至多10個,較佳地至多8個,更佳地至多5個,最佳地至多3個氨基酸被性質相似或相近的氨基酸所替換而形成多肽。在所述蛋白中,用性能相近或相似的氨基酸進行取代時,通常不會改變蛋白質的功能,在c末端和/或\末端添加一個或數個氨基酸通常也不會改變蛋白質的功能。這些保守性變異多肽最好根據表1進行氨基酸替換而產生。表1重組技術和植物改良本發明棉纖維長度相關基因的全長序列或其片段通常可以用pcr擴增法、重組法或人工合成的方法獲得。對于pcr擴增法,可根據本發明所公開的有關核苷酸序列,尤其是開放閱讀框序列來設計引物,并用市售的cdna庫或按本領域技術人員已知的常規方法所制備的cdna庫作為模板,擴增而得有關序列。當序列較長時,常常需要進行兩次或多次pcr擴增,然后再將各次擴增出的片段按正確次序拼接在一起。一旦獲得了有關的序列,就可以用重組法來大批量地獲得有關序列。這通常是將其克隆入載體,再轉入細胞,然后通過常規方法從增殖后的宿主細胞中分離得到有關序列。此外,還可用人工合成的方法來合成有關序列,尤其是片段長度較短時。通常,通過先合成多個小片段,然后再進行連接可獲得序列很長的片段。目前,已經可以完全通過化學合成來得到編碼本發明蛋白(或其片段,或其衍生物)的dna序列。然后可將該dna序列引入本領域中已知的各種現有的dna分子(或如載體)和細胞中。此外,還可通過化學合成將突變引入本發明蛋白序列中。本發明也涉及包含本發明的多核苷酸的載體,以及用本發明的載體或棉纖維長度相關多肽編碼序列經基因工程產生的宿主細胞,以及經重組技術產生本發明所述多肽的方法。通過常規的重組dna技術(science,1984;224:1431),可利用本發明的多聚核苷酸序列可用來表達或生產重組的棉纖維長度相關多肽。一般來說有以下步驟:(1)用本發明的多核苷酸(或變異體),或用含有該多核苷酸的重組表達載體轉化或轉導合適的宿主細胞;(2)在合適的培養基中培養的宿主細胞;(3)從培養基或細胞中分離、純化蛋白質。本發明的多核苷酸序列可插入到重組表達載體中。術語“重組表達載體”指本領域熟知的細菌質粒、噬菌體、酵母質粒、植物細胞病毒、哺乳動物細胞病毒或其他載體。總之,只要能在宿主體內復制和穩定,任何質粒和載體都可以用。表達載體的一個重要特征是通常含有復制起點、啟動子、標記基因和翻譯控制元件。本領域的技術人員熟知的方法能用于構建含本發明多核苷酸和合適的轉錄/翻譯控制信號的表達載體。這些方法包括體外重組dna技術、dna合成技術、 體內重組技術等。所述的dna序列可有效連接到表達載體中的適當啟動子上,以指導mrna合成。表達載體還包括翻譯起始用的核糖體結合位點和轉錄終止子。此外,表達載體優選地包含一個或多個選擇性標記基因,以提供用于選擇轉化的宿主細胞的表型性狀,如真核細胞培養用的二氫葉酸還原酶、新霉素抗性以及綠色熒光蛋白(gfp),或用于大腸桿菌的四環素或氨芐青霉素抗性。包含上述的適當dna序列以及適當啟動子或者控制序列的載體,可以用于轉化適當的宿主細胞,以使其能夠表達蛋白質。宿主細胞可以是原核細胞,如細菌細胞;或是低等真核細胞,如酵母細胞;或是高等真核細胞,如植物細胞(如農作物和林業植物的細胞)。代表性例子有:大腸桿菌,鏈霉菌屬、農桿菌;真菌細胞如酵母;植物細胞等。本發明的多核苷酸在高等真核細胞中表達時,如果在載體中插入增強子序列時將會使轉錄得到增強。增強子是dna的順式作用因子,通常大約有10到300個堿基對,作用于啟動子以增強基因的轉錄。本領域一般技術人員都清楚如何選擇適當的載體、啟動子、增強子和宿主細胞。用重組dna轉化宿主細胞可用本領域技術人員熟知的常規技術進行。當宿主為原核生物如大腸桿菌時,能吸收dna的感受態細胞可在指數生長期后收獲,用cacl2法處理,所用的步驟在本領域眾所周知。另一種方法是使用mgcl2。如果需要,轉化也可用電穿孔的方法進行。當宿主是真核生物,可選用如下的dna轉染方法:磷酸鈣共沉淀法,常規機械方法如顯微注射、電穿孔、脂質體包裝等。轉化植物也可使用農桿菌轉化或基因槍轉化等方法,例如葉盤法。對于轉化的植物細胞、組織或器官可以用常規方法再生成植株,從而獲得棉纖維長度變化的植物。獲得的轉化子可以用常規方法培養,表達本發明的基因所編碼的多肽。根據所用的宿主細胞,培養中所用的培養基可選自各種常規培養基。在適于宿主細胞生長的條件下進行培養。當宿主細胞生長到適當的細胞密度后,用合適的方法(如溫度轉換或化學誘導)誘導選擇的啟動子,將細胞再培養一段時間。在上面的方法中的重組多肽可在細胞內、或在細胞膜上表達、或分泌到細胞外。如果需要,可利用其物理的、化學的和其它特性通過各種分離方法分離和純化重組的蛋白。這些方法是本領域技術人員所熟知的。這些方法的例子包括但并不限于:常規的復性處理、用蛋白沉淀劑處理(鹽析方法)、離心、滲透破菌、超濾處理、超離心、分子篩層析(凝膠過濾)、吸附層析、離子交換層析、高效液相層析(hplc)和其它各種液相層析技術及這些方法的結合。重組的棉纖維長度相關多肽有多方面的用途。例如用于篩選具有調控棉纖維長度功能的化合物、多肽或其它配體。用表達的重組棉纖維長度相關多肽篩選多肽庫可用于尋找有價值的能抑制、或促進棉纖維長度相關多肽功能的多肽分子。另一方面,本發明還包括對棉纖維長度相關多肽具有特異性的多克隆抗體和單克隆抗體,尤其是單克隆抗體。本發明不僅包括完整的單克隆或多克隆抗體,而且還包括具有免疫活性的抗體片段、或嵌合抗體。本發明的抗體可以通過本領域內技術人員已知的各種技術進行制備。例如,純化的棉纖維長度相關多肽基因產物或者其具有抗原性的片段,可被施用于動物以誘導多克隆抗體的產生。本發明的各類抗體可以利用棉纖維長度相關基因產物的片段或功能區,通過常規免疫技術獲得。這些片段或功能區可以利用重組方法制備或利用多肽合成儀合成。與棉纖維長度相關基因產物的未修飾形式結合的抗體可以用原核細胞(例如e.coli)中生產的基因產物來免疫動物而產生;與翻譯后修飾形式結合的抗體(如糖基化或磷酸化的蛋白或多肽),可以用真核細胞(例如酵母或昆蟲細胞)中產生的基因產物來免疫動物而獲得。抗棉纖維長度相關多肽的抗體可用于檢測樣品中的棉纖維長度相關多肽。本發明還涉及定量和定位檢測棉纖維長度相關多肽水平的測試方法。這些試驗是本領域所熟知的。試驗中所檢測的棉纖維長度相關多肽水平,可用于解釋棉纖維長度相關多肽調控棉纖維長度的功能。一種檢測樣品中是否存在棉纖維長度相關多肽的方法是利用棉纖維長度相關多肽的特異性抗體進行檢測,它包括:將樣品與棉纖維長度相關多肽特異性抗體接觸;觀察是否形成抗體復合物,形成了抗體復合物就表示樣品中存在棉纖維長度相關多肽。本發明的多核苷酸的一部分或全部可作為探針固定在微陣列(microarray)或dna芯片(又稱為“基因芯片”)上,用于分析組織中基因的差異表達分析。用棉纖維長度相關多肽特異的引物進行rna-聚合酶鏈反應(rt-pcr)體外擴增也可檢測棉纖維長度相關多肽的轉錄產物。本發明的主要優點包括:(a)提供了棉纖維長度相關基因及其所編碼的多肽。(b)本發明的ghpre1基因可以促進纖維細胞伸長。(c)提供了一種有針對性地調控棉纖維長度的方法。(d)提供了一種改進棉纖維斷裂比強度的方法。(e)提供了一種改進棉纖維馬克隆值的方法。(f)提供了一種改進棉纖維紡紗均勻性的方法。(g)提供的ghpre1基因和方法同樣適用于其它植物的遺傳改良。下面結合具體實施例,進一步闡述本發明。應理解,這些實施例僅用于說明本發明而不用于限制本發明的范圍。下列實施例中未注明具體條件的實驗方法,通常按照常規條件,或按照制造廠商所建議的條件。除非另外說明,否則百分比和份數按重量計算。實施例1ghpre1cdna的分離棉花rna提取采用冷酚法。采集陸地棉r15(“晉棉r15”,購自山西省農科院棉花研究所)開花后6天的胚珠表面的纖維,在液氮中磨成粉末,轉移到50ml離心管中,加入8ml提取緩沖液(1mtris·hcl,50mmedta,1%sds,ph9.0)和等體積的水飽和酚:氯仿:異戊醇(25:24:1),震蕩混勻,冰上放置1h,每隔10min混勻一次。4℃,13000g離心20min。重復酚:氯仿:異戊醇抽提2~4次,最后用氯仿:異戊醇(24:1)抽提一次。取上清夜,加入1/2體積的高鹽溶液(0.8m檸檬酸鈉,1.2mnacl)和1/2體積異丙醇,混勻,-70℃放置1h。4℃,13000g離心20min,去上清液,沉淀溶于1mldepc處理水,4℃,13000g離心10min。上清液轉入1.5ml離心管,加入1/3體積的8mlicl和體積的naac,-20℃放置過夜。4℃,13000g離心20min。去上清液,用1ml70%乙醇清洗沉淀2次,室溫吹20min,溶于100~200μldepc處理水。根據ghpre1cdna的序列,合成專一引物(含酶切位點和保護堿基):ghpre1-bamhi-f5'-cgggatccatgtcaagcagaaggtggag-3'seqidno.:4ghpre1-kpni-r5'-ggggtaccttaattaaataagcttcgga-3'seqidno.:5以開花后6天的纖維總rna的反轉錄產物為模板做pcr反應,反應條件:94℃預變性5min;然后94℃變性30s,56℃復性30s,72℃延伸1min,共35個循環;最后72℃延伸10min。pcr產物電泳純化、回收后亞克隆到商業化載體pmd18-t上后經測序確認序列正確。實施例235s::ghpre1、rdl1::ghpre1和35s::dsghpre1等表達載體的構建和農桿菌轉化a.35s::ghpre1和rdl1::ghpre1植物表達載體構建首先對pcambia-2301植物表達載體進行改造,然后裝入目標基因,具體步驟如下:(1)在酶切位點hindiii-psti和saci-ecori間分別引入35s啟動子和nos終止子構建p35s::2301中間載體。(2)在酶切位點hindiii-psti和saci-ecori間分別引入prdl1啟動子和nos終止子構建prdl1::2301中間載體。(3)設計ghpre1全長cdna的pcr擴增引物,同時分別引入相應酶切位點用primestarhs高保真聚合酶擴增目標片段(見實施例1)。(4)核對讀碼框后,ghpre1片段經bamhii和kpni雙酶切,連到p35s::2301載體的相應位點中,形成35s::ghpre1植物表達載體,并經測序驗證序列正確無誤。(5)核對讀碼框后,ghpre1片段經bamhii和kpni雙酶切,連到prdl1::2301載體的相應位點中,形成rdl1::ghpre1植物表達載體,并經測序驗證序列正確無誤。b.35s::dsghpre1表達載體構建高保真酶擴增選ghpre1片段,正向片段引入saci和noti酶切位點,反向片段引入smai和xbai酶切位點,正向及反向序列分別克隆至含rtm內含子的pbsk載體上rtm序列兩端,用smai和saci酶切后替換35s::nos/p121上的gus基因,形成35s::dsghpre1植物表達載體,并經測序驗證序列正確無誤。rna干擾的引物序列:c.35s::ghpre1-venus表達載體構建(1)設計ghpre1全長cdna的pcr擴增引物,反向引物去除終止密碼子;同時分別引入相應酶切位點用primestarhs高保真聚合酶擴增目標片段。pcr引物序列如下:ghpre1-bamhi-f5'-cgggatccatgtcaagcagaaggtggag-3'seqidno.:10ghpre1-smai-r5'-tcccccgggattaaataagcttcggatgatg-3'seqidno.:11(2)核對讀碼框后,ghpre1片段經bamhii和smai雙酶切,連到p35s::2301載體的相應位點中,形成35s::ghpre1植物表達載體。(3)設計熒光蛋白venus全長cdna的pcr擴增引物,同時分別引入相應酶切位點用primestarhs高保真聚合酶擴增目標片段。pcr引物序列如下:(4)熒光蛋白venuspcr產物經smai和kpni雙酶切后連接到35s::ghpre1植物表達載體相應的酶切位點中,形成35s::ghpre1-venus植物表達載體。d.根癌農桿菌轉化根癌農桿菌的轉化采用凍融法。一個單菌落lba4404或gv3101(invitrogen),3mllb培養基(25μg/ml利福霉素rif和50μg/ml卡那霉素kan或慶大霉素gen),28℃,220rpm,過夜培養。2ml菌液,50mllb培養基(25μg/mlrif和50μg/mlgen),28℃,220rpm,培養到od600=0.5(約6h)。在冰上放置30min,4℃,5000g離心5min。重懸于10ml0.15mnacl。4℃,5000g離心5min。重懸于1ml20mmcacl2,50μl/管分裝,液氮速凍,-70℃保存感受態細胞。混合含目的基因雙元載體和50μl/管感受態細胞,在冰上放置30分鐘,液氮速凍1min。在37℃水浴中5min使菌液融化,加1mllb培養基,28℃,220rpm,培養2~4h。取50~100μl涂lb培養基平板(25μg/mlrif、50μg/mlgen和50μg/ml卡那霉素kan或潮霉素hyg),2天后挑單菌落為模板,并以目標基因引物進行pcr鑒定。將pcr鑒定為陽性的單克隆進行擴搖,進行下一步的植物轉化。實施例3植物轉化以及轉基因后代的篩選a.棉花的轉基因含rdl1::ghpre1和35s::dsghpre1載體質粒的農桿菌分別于添加卡那霉素50mg/l、利福平100mg/l、鏈霉素300mg/l的yeb細菌培養基上培養2~3天后,挑單菌落接種于含相同抗生素的yeb液體培養基中,于28℃、200rpm/min的搖床上懸浮培養過夜。菌液于4000rpm/min離心10min,沉淀用含葡萄糖30g/l和乙酰丁香酮100μmol/l的1/2ms液體培養基重新懸浮,調od600值為0.4~0.6左右,作為感染液備用。棉花r15種子(晉棉r15品種)經常規消毒后置于1/2ms0(1/2ms鹽+5g/l葡萄糖+7g/l瓊脂粉,ph6.0)培養基,在黑暗中萌發培養,5~7天后將無菌苗下胚軸切成1.0cm左右的切段作為轉化外植體備用。外植體在農桿菌菌液中浸泡感染15~20min,轉移到共培養培養基msb1(ms鹽+b5有機+30g/l葡萄糖+0.1mg/lkt+0.1mg/l2,4-d+2.2g/lgelrite,ph6.0)上,22℃暗培養2天后,將外植體轉移到培養基msb2(msb1+500mg/l頭孢霉素+80mg/l卡那霉素)上進行愈傷組織的誘導。外植體經過抗性愈傷組織的誘導、愈傷組織的增殖及胚性愈傷的誘導(培養基msb3:ms鹽+b5有機+30g/l葡萄糖+2.5g/lgelrite,ph6.0)、體細胞胚胎發生(培養基msb4:ms鹽+b5有機+30g/l葡萄糖+1.0g/l天門冬酰氨胺+2.0g/l谷氨酰胺+3.0g/lgelrite,ph6.0;ms鹽中kno3加倍,去除nh4no3),再生抗性試管苗。待試管苗長到3-4片真葉時,移栽到花盆中,放入人工氣候室生長。通過遺傳轉化共獲得5個不同的t0代rdl1::ghpre1轉基因棉花陽性株系, 以及7個不同的t0代35s::dsghpre1轉基因棉花陽性株系。后期經rt-pcr表達量檢測,篩選獲得2個rdl1::ghpre1轉基因棉花株系和3個35s::dsghpre1轉基因棉花株系用于后續分析。b.擬南芥的轉基因擬南芥植物的轉化采用花芽浸泡法(floraldip)(clough和bent,1998,plantj.16,735-743)。含35s::ghpre1、35s::ghpre1-venus載體質粒的農桿菌培養方法同上,菌液于4000rpm/min離心10min后,菌體重懸于500ml含0.02%silwetl-77的5%蔗糖溶液中。植株地上部分在菌液中浸泡5s,平放于塑料盆內,保濕,避光,16~24h。t0代種子在4℃春化2~4天,用20%漂水處理15min,無菌水清洗3~4遍。懸于0.5%的瓊脂糖(55℃),鋪在0.6%瓊脂的lb培養基(50μg/mlkan或hyg),22℃,連續光照,約一周后,將綠色抗性苗移栽到營養土(泥炭:蛭石:珍珠巖1:1:1)中生長。收取t1代種子在含卡那霉素的抗性板上萌發,篩選陽性苗并移栽到營養土中生長;收取t2代在含卡那霉素的抗性板上萌發,觀察分離比,篩選轉基因純合株系。選取純合株系的種子在垂直板上萌發,發現陽性植株的下胚軸和主根等部位具有顯著伸長的表型(圖3a)。選取有伸長表型的株系,提取rna并反轉錄檢測目標基因ghpre1的表達量(見實施例4),發現ghpre1的含量在陽性擬南芥苗中顯著積累(圖3b)。將陽性苗移栽到營養土中生長,3周后發現擬南芥葉柄等部位和對照相比也有顯著伸長的表型(圖3c)。實施例4轉基因植物的分子生物學鑒定a.pcr鑒定dna提取采用冷酚法。2g材料,在液氮中磨成粉末,轉移到50ml離心管中,加入8ml提取緩沖液(1mtris-hcl,50mmedta,1%sds,ph9.0)和等體積的水飽和酚:氯仿:異戊醇(25:24:1),震蕩混勻,冰上放置1h,每隔10min混勻一次。4℃,13000g離心20min。重復酚:氯仿:異戊醇抽提2~4次,最后用氯仿:異戊醇(24:1)抽提一次。取上清夜,加入1/2體積的高鹽溶液(0.8m檸檬酸鈉,1.2mnacl)和1/2體積異丙醇,混勻,-70℃放置1h。4℃,13000g離心20min,去上清液,用1ml70%乙醇清洗沉淀2次,室溫吹20min,沉淀溶于1ml無菌水。4℃,13000g離心10min。取上清液,加5~10μlrnase(10mg/ml),37℃消化30min。pcr鑒定采用nptii特異的引物:nptii-f:5'-ggagcaaggtgagatgacaggagatc-3'seqidno.:14nptii-r:5'-gattgtctgttgtgcccagtcatagc-3'seqidno.:15其反應條件為:94℃預變性5min;然后94℃預變性30sec,56℃復性30sec,72℃延伸1min,共35個循環;最后72℃延伸10min。擴增片段大小為680bp。rdl1::ghpre1和35s::dsghpre1轉基因棉花的pcr分子鑒定結果見圖4。b.熒光定量rt-pcr分析a.rt-pcr分析1μg的總rna,經oligo(dt)引物反轉錄,20μl反應液。42℃反應30分鐘,反轉錄第一鏈cdna,放置過夜。取0.5μl的反轉錄產物,25μlpcr反應體系檢測目的基因表達。pcr引物為ghpre1特異的引物:ghpre1-qrt-f:5'-gaaacctccttctgtattcctcc-3'seqidno.:16ghpre1-qrt-r:5'-aagggaagcatagtgtctcaaaat-3'seqidno.:17以棉花基因histone3(af024716)作為內標,校正rt-pcr反應的模板量。pcr反應條件為:94℃預變性5分鐘;然后94℃變性30秒鐘,56℃復性30秒鐘,72℃延伸1分鐘,共30個循環;最后72℃延伸10分鐘。b.熒光定量rt-pcr分析定量rt-pcr檢測采用嵌合熒光法(premixextaqtmii(perfectrealtime),takara,drr041a)。反應體系如下:以棉花基因histone3(af024716)作為內標。數據分析采用realplexv2.0(購自德國漢堡eppendorf,nsw)。實驗重復三次,取各組數據的平均值和方差,繪制圖表。通過熒光定量pcr分析了ghpre1在棉花根、莖、葉和胚珠等不同組織的表達特征,發現該基因在棉花胚珠中有較高的表達量(圖2b);進一步分析了不同發育時期棉纖維細胞中ghpre1的表達水平,發現其在開花后3-12天的纖維細胞中高表達,特別是在開花后6天左右的纖維細胞中表達量最高(圖2a)。ghpre1在伸長期的纖維細胞中特異高表達,暗示其在棉纖維的伸長發育過程中行使重要功能。亞細胞定位顯示ghpre1-venus融合蛋白定位于細胞核和細胞質(圖2c)。實施例5轉基因棉花的性狀分析35s::dsghpre1、rdl1::ghpre1轉基因棉花和野生型r15在上海和海南農場種植,常規田間管理。在棉花成熟吐絮期,分別單株收取成熟棉桃,盡量保持收取部位的一致性,隨機取一定數量的種子將其纖維梳平并測量其纖維長度,進行統計分析。結果表明,rdl1::ghpre1轉基因棉花纖維長度顯著增加(圖5a,b),轉基因棉花纖維平均長度達到32mm,部分纖維長度達到34mm以上(對照受體棉纖維長度為28mm左右;基因表達檢測顯示在t2代轉基因棉纖維中ghpre1的含量顯著增加(圖5c)。而35s::dsghpre1抑制ghpre1表達的轉基因植株的基因表達檢測顯示棉纖維中ghpre1的含量被顯著下調(圖6c),纖維長度表現出顯著變短的表型(圖6a,b)。將rdl1::ghpre1轉基因棉花纖維送往中國農業科學院棉花研究所的“農業部棉花品質監督檢驗測試中心”進行進一步的品質檢測。檢測結果如圖8所示,rdl1::ghpre1轉基因棉花纖維長度顯著增加。轉基因棉花上半部平均長度為32.1-33.9mm,對照受體棉纖維長度為29.1-29.8mm。并且,rdl1::ghpre1轉基因棉花纖維的斷裂比強度顯著優于對照;馬克隆值處于國家b2級標準且優于對照;紡紗一致性系數顯著高于對照,表明rdl1::ghpre1轉基因棉花纖維的成紗更強力,可紡性更好。圖8的棉花纖維檢測結果報告書由中國農業科學院棉花研究所的“農業部棉花品質監督檢驗測試中心”出具。ox-2-1/2或ox-7-2表示rdl1::ghpre1同一轉基因株系在不同大田區域的重復,wt表示野生型對照。“*”表示rdl1::ghpre1轉基因棉花的纖維品質顯著優于對照組(wt)。依據中華人民共和國農業部第2244號公告,纖維品質各個指標的定義如下:(1)上半部平均長度(len)是棉纖維長度分布中纖維平均長度以上的所有纖維的長度平均值,單位毫米。(2)整齊度指數(unf)是平均長度和上半部分平均長度的比值。(3)斷裂比強度(str)是棉花纖維試樣受到拉伸直至斷裂時,所顯示出來每單位線密度所受的力,單位cn/tex。國家標準規定斷裂比強度分五檔:s1代表很強,比強度大于等于31cn/tex;s2代表強,29.0~30.9cn/tex;s3代表中等,26.0~28.9cn/tex;s4代表差,24.0~25.9cn/tex;s5代表很差,比強度小于24.0cn/tex。(4)馬克隆值(mic)是棉花纖維細度與成熟度的綜合指標。馬克隆值越大,表明棉花纖維越粗和成熟度越好,反之,棉花纖維越細和成熟度較差。按 國家棉花標準馬克隆值分為:a級范圍為3.7-4.2;b1級范圍為3.5-3.6;b2級范圍為4.3-4.9;c1級范圍為3.4及以下;c2級范圍為5.0及以上。其中,a級最優,b1、b2次之,c1、c2較差。(5)伸長率(elong)是棉花纖維試樣受到拉伸直至斷裂時,纖維的絕對伸長長度與纖維正常伸展長度的比值。表征棉纖維抵抗拉伸的能力。(6)反射率(rd)是棉花對光的反射程度。反射率高,表明棉花纖維成熟較好,色澤好。(7)黃度(+b)是棉花白度差別的物理量。一般來說,黃度低,品級高。(8)紡紗一致性系數(sci)是大容量纖維測試儀(highvolumeinstrument,hvi)提供一個多重回歸經驗性公式,它能反映纖維的可紡性和估算成紗強力。公式中共涉及測試項目中纖維斷裂比強度、馬克隆值、上半部平均長度、長度整齊度、反射率、黃色深度6個參數,其中前4個為主要參數,影響較大。一般計算公式如下:sci=-414.67+2.9×str-9.32×mic+49.17×len+4.74×unf+0.65×rd+0.36×(+b)一般情況下,sci值越大,成紗強力和可紡性越好。實施例6轉基因擬南芥的性狀分析35s::ghpre1和35s::ghpre1-venus轉基因載體通過農桿菌介導的花芽浸泡法轉入擬南芥,通過抗性篩選和rt-pcr的方法得到純合的轉基因陽性植株。取35s::ghpre1和35s::ghpre1-venus轉基因種子和同時收取的野生型種子在ms培養基上萌發,觀察植物表型。結果表明,35s::ghpre1轉基因擬南芥幼苗下胚軸和主根等部位都有顯著增長的表型,并且ghpre1表達量越高,增長的表型越明顯(圖3a,b)。對萌發后3周的小苗進行觀察,發現ghpre1高表達的擬南芥葉柄較對照組長得多(圖3c)。35s::ghpre1-venus轉基因擬南芥與35s::ghpre1轉基因擬南芥具有相同的表型變化(圖3c),說明棉花ghpre1在異種植物中的高表達也能促進植物伸長生長。對比例1棉纖維發育是一個復雜的多基因控制的動態過程。至今已有一些棉纖維功能基因的研究報道,大多數轉基因棉花的結果是可以抑制棉纖維伸長,但通過轉基因技術成功提高纖維品質的報道并不多。e6基因是最早分離得到的棉纖維高表達基因,通過rnai技術下調e6基因表達(僅為野生型水平的2-40%),卻未 對相應的轉基因植株及其纖維發育造成影響(johnme,andkellerg.(1996).structuralcharacterizationofgenescorrespondingtocottonfibermrna,e6:reducede6proteinintransgenicplantsbyantisensegene.plantmolecularbiology,1996,30(2):297-306)。申請人分離了若干棉纖維特異高表達的基因,如gamyb2和gahox1,功能分析表明它們與擬南芥表皮毛發育的調控因子gl1和gl2具有一致的功能(wangs,wangjw,etal.2004.controlofplanttrichomedevelopmentbyacottonfibermybgene.plantcell16(9):2323-2334.和guanxy,liqj,etal.(2008).thehd-zipivgenegahox1fromcottonisafunctionalhomologueofthearabidopsisglabra2.physiologiaplantarum134:174-182.),可能對棉纖維的起始和伸長具有重要功能。但轉基因實驗結果表明,抑制這些基因的表達可引起棉纖維變短,但是過表達這些基因并不能使纖維增長。申請人近期分離了一個新的棉纖維高表達的基因ghacy,該基因在開花后3-12天快速伸長的纖維細胞中高表達(圖7a)。通過農桿菌轉化獲得了ghacy抑制與高表達的轉基因棉花(方法同實施例3),對t2轉基因棉花的纖維進行長度分析,發現ghacy表達受抑制的棉纖長度有輕微變短的表型(圖7b,c);過表達ghacy的轉基因棉花纖維長度則并無顯著變化(圖7d,e)。實施例與對比例1的比較說明本發明提供的ghpre1基因及其轉基因構建方式可特異地實現對植物生長發育的調控,以獲得在育種上有利用價值的材料。在本發明提及的所有文獻都在本申請中引用作為參考,就如同每一篇文獻被單獨引用作為參考那樣。此外應理解,在閱讀了本發明的上述講授內容之后,本領域技術人員可以對本發明作各種改動或修改,這些等價形式同樣落于本申請所附權利要求書所限定的范圍。當前第1頁12
相關技術
網友詢問留言
已有0條留言
- 還沒有人留言評論。精彩留言會獲得點贊!
1